Under normal physiological conditions, brain pericytes generally do not undergo cell death. Figure 6 illustrates this phenomenon, with yellow indicating the presence of vital brain pericytes; brain pericytes show no staining with PI, indicating their viability. To further investigate whether pericytes remain attached to the microvasculature following cell death, methods were employed in a SAH rat model, and subsequent imaging was conducted.
Methods for imaging both vital and non-vital brain pericytes in brain slices after SAH have been developed. As depicted in Figure 7, vital brain pericytes (blue arrows) are located within the microvasculature, while non-vital brain pericytes are represented by white arrows. This simultaneous visualization allows for the identification of both vital and non-vital brain pericytes within brain slices. Furthermore, it was observed that PI-labeled non-vital brain pericytes remained attached to the entire microvasculature.
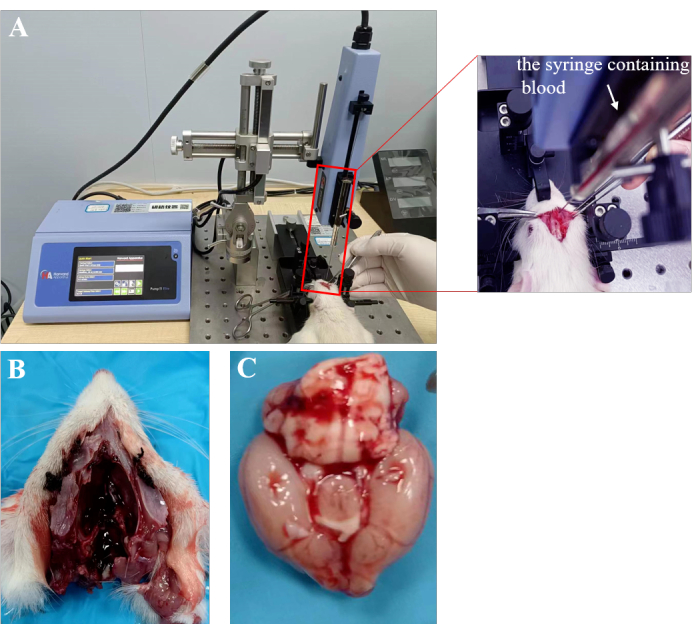
Figure 1: SAH model. (A) The rat's head was firmly secured to the stereotaxic apparatus to ensure stability. The SAH model was induced by carefully inserting a stereotaxic needle into the suprasellar cistern. (B,C) In the SAH model, blood enters the subarachnoid space within the skull, affecting the brain. The cerebral hemispheres fill with blood approximately 24 h after SAH. Please click here to view a larger version of this figure.
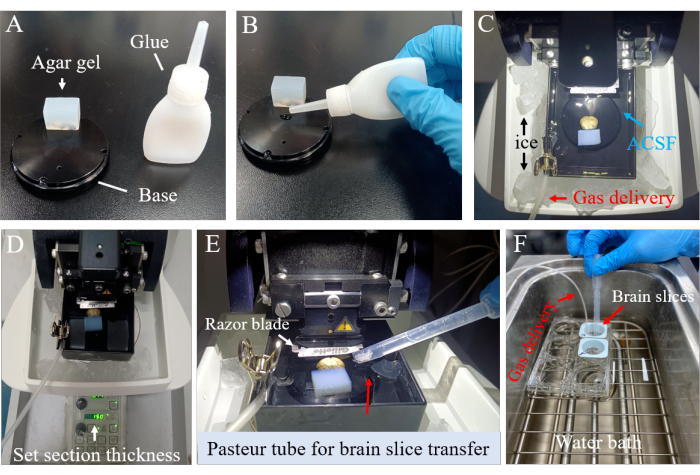
Figure 2: Acute whole brain slice preparation. (A) The bottom chamber was coated with agarose glue for fixation. (B) The glue was meticulously applied to the bottom chamber to adhere firmly to the brain. (C) The surrounding tank was filled with ice-cold ACSF to maintain temperature. (D) The desired section thickness was carefully defined. (E,F). The brain slices were meticulously transferred to a 6-well plate. Please click here to view a larger version of this figure.
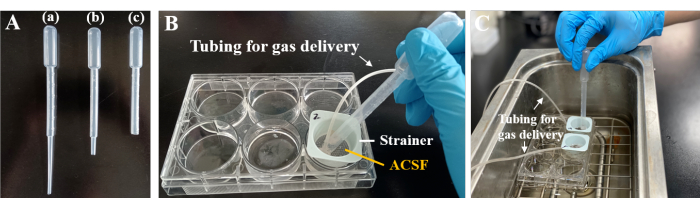
Figure 3: Transfer pipettes and dye-loading in a 6-well plate. (A) Acute brain slices were transferred using plastic Pasteur pipettes (3 mL). The length of the pipette used in (a) was 18.2 cm. For (b) and (c), the fine tip of the plastic Pasteur pipette was carefully trimmed to prevent any potential damage to the brain slices during transfer. The 6-well plates were efficiently transitioned from (B) to (C) for fluorescence staining in a 37 °C water bath. (B) One well of the 6-well plate accommodated a small nylon mesh strainer, with fine tubing positioned for gas delivery. Please click here to view a larger version of this figure.
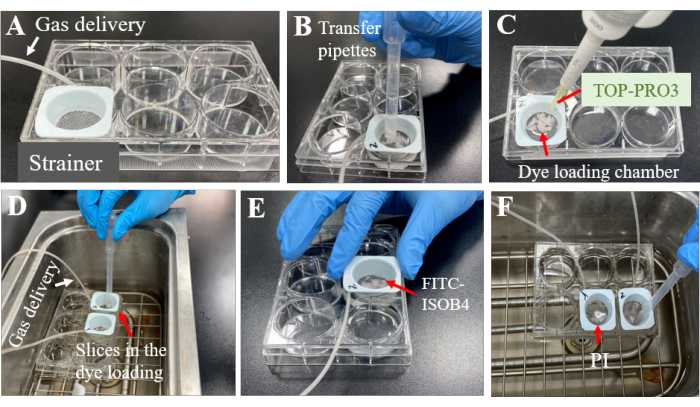
Figure 4: Incubation of acute brain slices. A systematic procedure was meticulously followed to label pericytes with TO-PRO-3 in the acute brain slices. (A) Initially, one well of a six-well plate (12 × 8 cm) was filled with 10 mL of ACSF, ensuring proper aeration by bubbling the solution using fine tubing connected to a mixture of 95% O2 and 5% CO2. (B) Subsequently, the brain slices were meticulously transferred to the six-well plate. (C) 10 µL of the TO-PRO-3 stock solution was introduced into the dye-loading chamber, gently agitating to facilitate dye dissolution (D). (E,F) To further characterize the brain slices, staining with IB4 (1 µM in ACSF) and PI (1 µM in ACSF) was conducted. These sequential steps enabled the successful labeling of pericytes with TO-PRO-3 in acute brain slices, thus facilitating subsequent analysis and examination of the labeled brain pericytes. Please click here to view a larger version of this figure.

Figure 5: Imaging of vital and non-vital brain pericytes in brain slices. (A) The solution is equilibrated with a mixture of 95% O2 and 5% CO2 delivered through the tubing. (B–E) Setup for high-resolution imaging of TO-PRO-3-labeled, IB4-labeled endothelial, and PI-labeled cells in acute brain slices. (E) Confocal microscopy. Please click here to view a larger version of this figure.
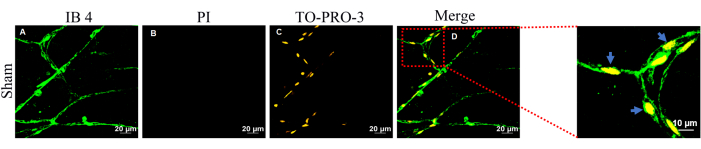
Figure 6: Image of vital brain pericytes in brain slices. (A) Cerebral microvasculature was labeled with IB4 (green). (B) Non-vital cells were labeled with PI. (C) Vital pericytes were labeled with TO-PRO-3 (yellow). (D) The merged image is displayed. Please click here to view a larger version of this figure.
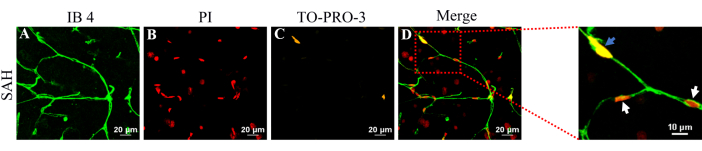
Figure 7: Representative image of vital and non-vital brain pericytes in a brain slice after SAH. (A) Cerebral microvasculature was labeled with IB4 (green). (B) Non-vital cells were labeled with PI (red). (C) Vital pericytes were labeled with TO-PRO-3 (yellow). (D) The merged image is displayed. Blue arrows indicate examples of vital brain pericytes, while white arrows indicate examples of non-vital brain pericytes. Notably, the nuclei of pericytes do not protrude above the microvascular surface. Please click here to view a larger version of this figure.
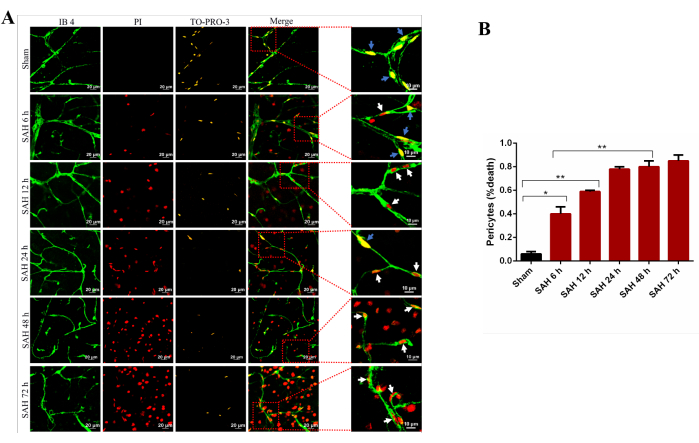
Figure 8: Positive correlation between the number of non-vital pericytes and time after SAH. High-resolution confocal imaging was employed to capture brain pericytes at various time points post SAH. Noticeable cell death of brain pericytes commenced at 6 h after SAH, as indicated by white arrows in (A). Subsequently, there was a substantial increase in pericyte death from the 6 h mark onward. The count of non-vital pericytes displayed a positive correlation with the time elapsed after SAH, as depicted in (B). Please click here to view a larger version of this figure.
