All experiments strictly followed the ethical guidelines set by the institutional review board at the University of Brasilia (process 13364819.0.0000.5558), and samples were identified by codes to ensure donor anonymity. The cells were obtained from normal healthy male donors aged 18-35 years, who signed the informed consent and met the following eligibility criteria: non-smokers/vapers, no chronic health conditions, and no history of inflammatory conditions in the last 14 days.
1. Blood collection
- Aseptically place 0.3 mL of 5,000 IU/mL heparin (see Table of Materials) in a sterile 20 mL syringe to heparinize it.
- Apply a venous tourniquet about 4 in above the puncture site and identify the median cubital or cephalic vein for venipuncture.
NOTE: Ensure that the total tourniquet time does not exceed 1 min. - Clean the puncture site with 70% alcohol and perform the venipuncture.
- Gently invert the syringe three or four times after collecting the blood to mix the blood and heparin properly.
2. Neutrophil isolation
NOTE: Polymorphonuclear leukocytes (PMNs) are isolated through density gradient centrifugation followed by hypotonic lysis of the remaining red blood cells (RBC), as previously described11 with some changes. This method is not mandatory to perform the screening assays, and can be replaced as long as the chosen method results in a viability of >97%, priming or activation of <3% of the PMNs, and yields enough cells for all assays, replicates, and conditions. Performing these steps under aseptic conditions and using endotoxin-free solutions are mandatory to avoid cell activation.
- Make 12 mL dilutions of 60% and 70% separation media (commercially available; see Table of Materials) in 50 mL conical tubes.
- Prepare the gradient from bottom to top by adding 4 mL at a time of the 60% dilution over the 70% dilution, using a 5 mL pipette. Do this gently to prevent mixing the interface.
- Carefully layer 12 mL of heparinized blood on top of the density gradient. Centrifuge at 200 x g for 15 min at room temperature.
NOTE: From this step onward, until PMN activation, all the reagents and tubes used must be kept in a cooler filled with ice. - Discard the plasma/mononuclear cell layer, then gently transfer the layer above the erythrocyte pellet into two 15 mL conical tubes with approximately 7.5 mL in each. Make up the tube volume with Hank's balanced salt solution (HBSS; see Table of Materials).
- Centrifuge at 300 x g for 5 min at 19 °C.
- Wash the cell pellet with HBSS.
- Discard the supernatant by pouring the tube and gently resuspend the pellet in 7 mL of HBSS.
- Centrifuge at 300 x g for 5 min at 19 °C to remove all the separation media.
- Perform hypotonic lysis of the remaining RBCs.
- Discard the supernatant and combine the pellets in a single tube.
- Resuspend the RBC/PMN pellet in 3 mL of sterile H2O and add 3 mL of HBSS (2x) within 25 s to restore the osmolarity. Then, centrifuge at 300 x g for 5 min at 19 °C.
- Repeat steps 2.8.1 and 2.8.2 for a white, erythrocyte-free pellet.
NOTE: The supernatant must be removed as soon as possible to minimize the contact of neutrophils with RBC breakdown products. Alternatively, the second hypotonic lysis can be replaced by gently resuspending the residual RBC layer and removing all the supernatant, as the remaining RBCs will sediment above the PMN pellet.
- Discard the supernatant by pouring the tube, gently resuspend the PMNs in the remaining buffer, and transfer them to an ice-cold microtube.
NOTE: Ensure to annotate the volume while transferring the resuspended cells with a micropipette. - Transfer 3 x 1 µL of the cell suspension to a clean glass slide (three wells of 1 µL each) and stain with fast panoptic (see Table of Materials) for morphology and purity evaluation12.
- To stain with fast panoptic, immerse the slide five times in panoptic fixative n° 1, six times in eosin n° 2, and twice in hematoxylin n° 3, with each immersion lasting 1 s.
- Gently wash the glass slide with distilled water.
- Allow to drain and air-dry.
- Observe under a microscope and count 300 random cells in each well, thus differentiating the neutrophils from other granulocytes.
- Transfer 1 µL of the cell suspension to 49 µL of 0.2% trypan blue dye13 and count the cells using a Neubauer chamber, distinguishing between dead and viable cells.
- Adjust the cell concentration to 6,667 cells/µL using a solution of 50% autologous plasma and 50% HBSS supplemented with calcium and magnesium. Divide the 6,667 cells/µL suspension evenly among the microtubes corresponding to the conditions to be tested, including the negative control.
NOTE: Any cell concentration similar to the circulating neutrophils in the model organism can be used, but it is important to use the same cell concentration in all experiments for reproducibility.
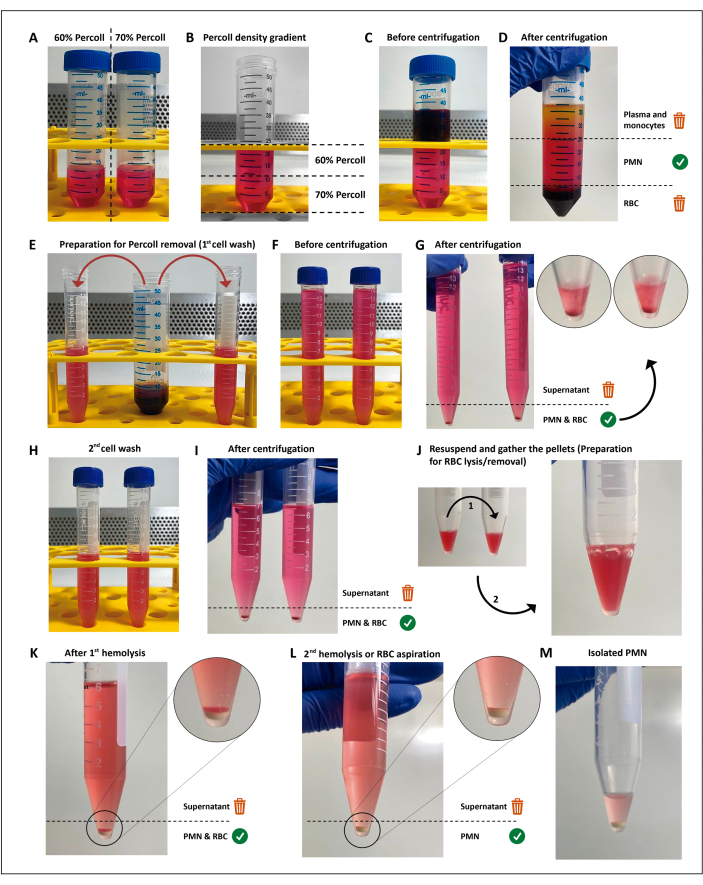
Figure 1: The neutrophil isolation protocol. Two concentrations of the separation media (percoll) (A) are stacked (B), then the blood is layered on top of the separation gradient (C). After centrifugation, the PMN is in the central layer (D), which is divided into two 15 mL tubes (E). The cell suspension is washed twice in HBSS and centrifuged (G–I) to remove the media, then the cells are resuspended, and residual RBCs are submitted to two rounds of hypotonic lysis (J–M). Please click here to view a larger version of this figure.
3. Preparation for neutrophil activation
- In 1.5 mL microtubes, prepare an activation system for each condition so that the final cell concentration is 6,600 cells/µL. For instance, to test the effects of 100 nM fMLP (N-formyl-methionyl-leucyl-phenylalanine; see Table of Materials), add 5 µL of 10 µM fMLP to 495 µL of the 6,667 cell/µL suspension. For the negative (unstimulated) control, add HBSS containing Ca2+ and Mg2+.
NOTE: To demonstrate this methodology, the following final concentrations of the stimuli were used: 100 nM fMLP, 16 µM of fallaxin, a naturally occurring antimicrobial peptide14, and 100 nM PMA (phorbol 12-myristate 13-acetate) (see Table of Materials). - Incubate at 37 °C without rotation.
NOTE: All aliquots for functional assays are taken from this cell suspension, hereafter referred to as the activation system.
4. Nitrotetrazolium blue chloride (NBT) assay for evaluating ROS production
- Preparation of NBT working solution: For each experimental condition, prepare an NBT (see Table of Materials) working solution of 6 mM using the following steps:
- Dissolve 0.0005 g of NBT in 10 µL of dimethyl sulfoxide (DMSO) and vortex for at least 15 min.
- Add 90 µL of HBSS Ca2+Mg2+ and vortex up to 2 min.
NOTE: All steps involving NBT must be performed in the dark.
- Perform NBT slide test.
- After 20 min of cell activation, gently mix the cell suspension and carefully transfer 2 µL of PMN to a clean glass slide. Incubate for 20 min in a humidified chamber at 37 °C.
NOTE: Do not spread the cell suspension too much over the slide; otherwise it may dry out before incubation. - Add 1 µL of the NBT working solution over the cells and further incubate for 20 min protected from light.
- Dry the slide with hot air and fix it with a drop of methanol in each well for 1 min. Stain with 0.03% safranin (see Table of Materials) for 1 min.
- Gently wash the glass slide with distilled water.
- Allow the slide to air-dry and observe under a microscope.
- Count 100 random cells in each well, differentiating neutrophils with and without formazan deposits.
- After 20 min of cell activation, gently mix the cell suspension and carefully transfer 2 µL of PMN to a clean glass slide. Incubate for 20 min in a humidified chamber at 37 °C.
- Perform NBT spectrophotometry assay.
- After 40 min of cell activation, gently mix the cell suspension and transfer 90 µL of PMNs from the activation system to a clean microtube. Then, carefully add 20 µL of the 6 mM NBT solution. Incubate in the dark for 20 min at 37 °C.
- Add 100 µL of 10% Sodium dodecyl sulfate (SDS; see Table of Materials) and vortex.
- Sonicate using a tip-sonicator at an amplitude of 60%, five cycles of 15 s each with 15 s intervals. Centrifuge at 12,000 x g for 5 min.
- Transfer 60 µL of the supernatant to a clear-bottom 96-well plate and measure the absorbance of the formazan product at 570 nm.
5. Phagocytosis assay
- Prepare a 33,000 yeasts/µL suspension for each condition, as described below:
- Add approximately 0.75 mg of dry yeast (Saccharomyces cerevisiae; see Table of Materials) to 200 µL of HBSS Ca2+Mg2+ and incubate in a thermomixer at 100 °C with 500 rpm for at least 15 min.
- Homogenize the mixture by vortexing and transfer 5 µL of yeast suspension to 45 µL of 0.2% trypan blue dye. Count the yeasts using a Neubauer chamber.
- Adjust the concentration of the initial suspension to 33,000 yeast cells/µL using HBSS Ca2+Mg2+. Keep the suspension on ice until use.
- After 20 min of cell activation, gently mix the cell suspension and transfer 5 µL of the activation system to 5 µL of the 33,000 yeast/µL Saccharomyces cerevisiae suspension in a new sterile microtube.
NOTE: The neutrophil to yeast ratio is 1:5 (PMN:yeast). - Immediately transfer 6 µL of the PMN/yeast suspension into three wells of a clean glass slide (2 µL each) and incubate the slide in a humidified chamber for 40 min.
NOTE: Do not spread the cell suspension too much over the slide; otherwise, it may dry out before incubation. - Dry the slide under hot air and stain with fast panoptic, as described in step 2.9 above.
NOTE: The third step of the fast panoptic staining is critical to the microscopy analysis of the slide. Staining the slide for ≥3 s at this step can render it unfit for analysis, since it will be difficult to differentiate yeast from neutrophil nuclear lobes. - Observe the slides under the microscope, counting 100 random neutrophils of each well and discriminating between PMNs positive and negative for phagocytosis.
NOTE: At least one yeast particle within or in direct contact with the PMN cellular membrane indicates a PMN positive for phagocytosis. If interested in the yeast/neutrophil ratio, count the number of yeast particles engulfed as well.
6. Real-time PMN chemotaxis assay
NOTE: The migration assay is performed similarly to the protocol described previoulsy15, with the following adaptations:
- Prepare the chemotactic gradient by adding 160 µL of chemoattractant (e.g., fMLP, IL-8, C5, or LTB4; see Table of Materials) to the lower chamber of an impedance-based real time cell analyzer (RTCA) plate. For negative controls and blanks, add 160 µL of HBSS Ca2+Mg2+.
- Attach the upper chamber and add 25 µL of HBSS Ca2+Mg2+. Incubate at room temperature for at least 1 h to form the chemotactic gradient.
- After 60 min of cell activation, gently mix the cell suspension and place 60 µL of cell suspension in the upper chamber. Add 60 µL of HBSS Ca2+Mg2+ to the blank.
- Place the RTCA plate and program the RTCA software to measure the cell index (CI) every 60 s for 2 h.
NOTE: The RTCA plates can be washed for reuse as previously described16. In summary, wash the RTCA chambers and electrodes with phosphate-buferred saline (PBS) three times, then with type I ultrapure water twice. Incubate the lower and upper chamber with 0.25% trypsin 0.53 mM ethylenediaminetetraacetic acid (EDTA) for 40 min. Wash with ultrapure water three times.
7. NET suggestive assay
- After 10 min of cell activation, gently mix the cell suspension and transfer 4 µL of the PMNs from each activation system under evaluation, divided into two wells of a clean glass slide. Incubate in a humidified chamber at 37 °C for 30 min.
- Add 1 µL of DNAse I to one of the wells and incubate for 20 min at 37 °C (in a wet chamber).
- Dry the slide and stain with fast panoptic, as previously described in step 2.9 of the "neutrophil isolation" section.
- Evaluate the slides under a microscope.
NOTE: Look for any indication of NET release, characterized by the presence of web-like structures. Once identified, confirm if the DNAse I treatment was capable of removing such structures. This assay is suggestive of NET formation, as additional tests are required to confirm their presence.
The density-based isolation method used in this study (Figure 1) met the criteria for the proposed experiments. Neutrophil parameters obtained from this method included viability ≥98%, purity ≥94%, and cell yield ≥1.5 x 107, with no activation detectable by the screening tests. Two relevant steps in the isolation of PMNs are anticoagulation and RBC removal. Keeping the anticoagulated blood tube or syringe at a gentle rocking before layering over the density gradient and choosing an RBC removal method to prevent both activation and contamination can influence the experiment yield and reproducibility.
To assess the functional overview of neutrophil function, a workflow was developed that allowed the proposed screening assays to be performed with at least two researchers in an average time of 4 h, with 1 h of activation and 2 h of real-time migration monitoring, as shown in Figure 2.
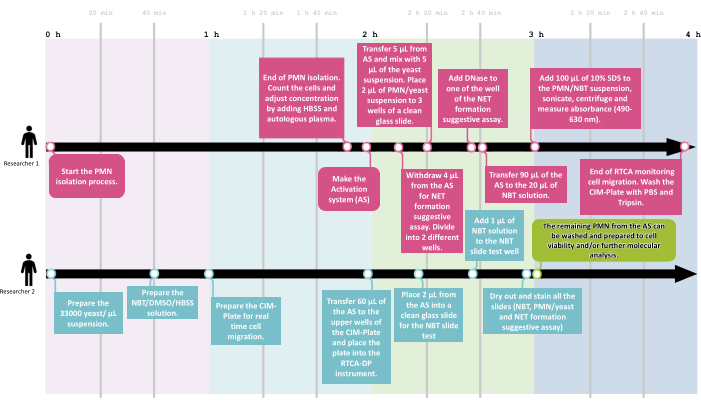
Figure 2: The NeutroFun Screen workflow. To assess the panel of functional responses triggered by specific conditions under evaluation, the NeutroFun Screen workflow includes the participation of at least two researchers. Researcher 1 (R1) starts the PMN isolation process, followed by cell concentration adjustment and activation system incubation, while in parallel, researcher 2 (R2) prepares the materials to carry out the next assays, which are divided between the two: R1 performs phagocytosis, NET, and spectrophotometric NBT assays; R2 performs real-time migration and NBT slide tests. Please click here to view a larger version of this figure.
The classic NBT assay17 was optimized for both slide and spectrophotometric assessment of the formazan formation resulting from the reaction of NBT with the ROS generated by PMNs. The spectrophotometric assay allowed for relative quantification among conditions, while the microscopy assay was useful for descriptive and semi-quantitative analysis, evaluating the regularity of crystal distribution, morphology, and number of cells containing formazan. The results of the spectrophotometric assay are shown in Figure 3A, indicating that 100 nM PMA induced elevated ROS production (in an average ratio of 3:2:1) compared to fMLP and control groups. The NBT slide test results (Figure 3B) corroborated the spectrophotometric results, also showing PMA as the most intense ROS production-inducing stimulus compared to fMLP (Figure 3C), as previously demonstrated by the direct measurement of ROS by cytometry18. Regarding cell counting, the number of neutrophils containing formazan crystals showed a ratio of 7:4:1 in the PMA:fMLP:control conditions, also revealing different formazan distribution patterns between fMLP and PMA-activated PMNs; fMLP treatment resulted in intensively activated individual cells, while PMA induced the formation of formazan crystals scattered throughout the cytoplasm in the majority of PMNs. Furthermore, a few morphological characteristics that differentiated the conditions were observed, such as expressive cell aggregation after PMA treatment. The absence of such typical activation characteristics and low levels of formazan on the control group indicated that the resting state was not significantly disturbed. Therefore, the NBT assay proved to be a simple, inexpensive, and reliable test for neutrophil ROS production that can be used to overview the capacity and intensity of a given stimulus to induce ROS production.
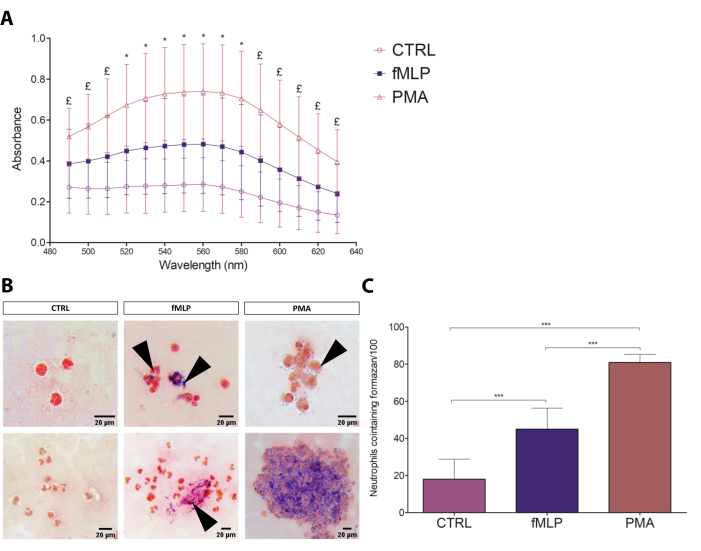
Figure 3: NBT test. (A) The spectrophotometric assessment of neutrophil respiratory burst to different conditions (negative control: 100 nM fMLP and 100 nM PMA) by formazan absorbance (490-630 nm). Data from four donors are presented as mean ± standard deviation (SD). * = all groups are different from each other (p≤ 0.05); £ = The comparisons CTRL versus PMA and fMLP versus PMA show significant differences (p≤ 0.05). (B) The slide test can be analyzed qualitatively, characterizing the intensity of formazan deposits, its location within the cell, and the cell aggregation. Black arrows point to formazan crystals. Scale bars: 20 µm. (C) Number of PMNs containing formazan, counted by optical microscopy. Data from eight donors presented as mean ± SD. *** p < 0.0001, Student's t-test. Please click here to view a larger version of this figure.
The phagocytosis slides were used to determine the phagocytosis rate as a percentage of neutrophils performing phagocytosis, as shown in Figure 4. Although both 100 nM fMLP and 16 µM antimicrobial peptide induced an apparently increased yeast engulfment, the difference was not statistically significant until the dual stimulation, which significantly enhanced the phagocytosis compared to the control group and might suggest a response upon dual stimulation.
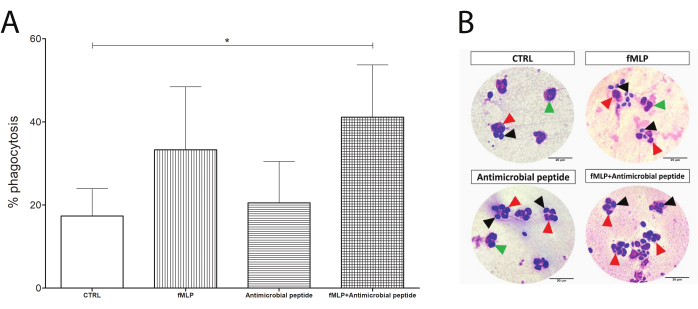
Figure 4: Phagocytosis test. Neutrophils were evaluated by their phagocytosis capacity (counting of neutrophils containing yeast). Although the exposure to 100 nM fMLP or a 16 µM antimicrobial peptide prior to yeast incubation enhanced the phagocytosis compared to the control group, (A) the increase was not statistically significant. Interestingly, an initial incubation with fMLP, followed by addition of the antimicrobial peptide, resulted in significantly enhanced phagocytosis in human neutrophils (asterisk between CTRL and the fMLP+ antimicrobial peptide). (B) Black arrows show yeast cells, green arrows show neutrophils alone, and red arrows point to phagocytosing neutrophils. Data from five donors presented as mean ± SD. Scale bars: 20 µm. Please click here to view a larger version of this figure.
An example of real-time cell migration results is shown in Figure 5, exhibiting the well-known chemotactic effect of 100 nM fMLP on neutrophils. The cell migration kinetics was adjusted to the Gompertz model, enabling the comparison of its parameters.
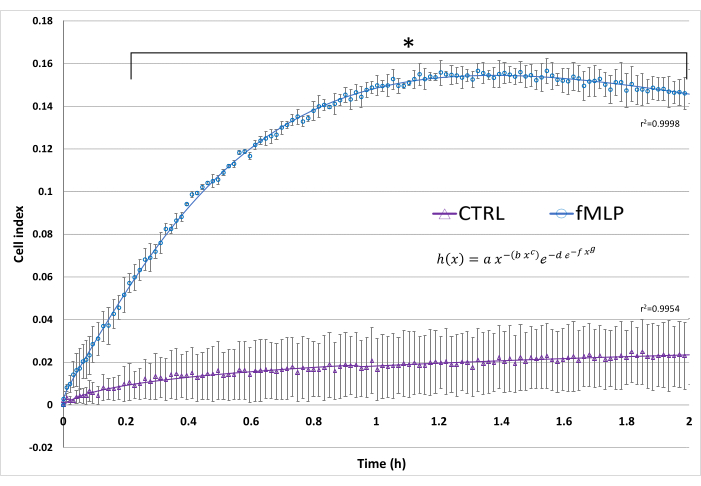
Figure 5: Real-time cell migration. The cell index reflects the electrical impedance resulting from cell migration. In this assay, fMLP was used as a positive control of neutrophil migration by adding 100 nM fMLP to the lower chamber. A negative control (CTRL) contained only HBSS in the lower chamber. The superimposed curves represent the curve fitting to the Gompertz model, modified to best fit ascending and descending slopes. Data from three donors, presented as mean ± SD.* p < 0.05, Student's t-test. Please click here to view a larger version of this figure.
Finally, a simple optical microscopy test suggestive of the presence of NET is presented, in which the PMNs are stained with fast panoptic. It is possible to observe that for some specific NET-inducing stimuli, such as PMA or some antimicrobial peptides, even after a short activation time (1 h) it is possible to observe filamentous web-like structures near the cells, while this is not possible in other conditions like the negative control and fMLP (Figure 6). To better support this claim, DNase I was added after 40 min of cell activation with 100 nm PMA or an antimicrobial peptide, removing the NET-like structures (Figure 6). Although this method is not suitable for NET characterization and can be influenced by artifacts, it is a cheap, simple, and interesting starting point to justify further investments in analyzing NETs, especially in contexts wherein the effects of the stimuli are unknown or poorly described.
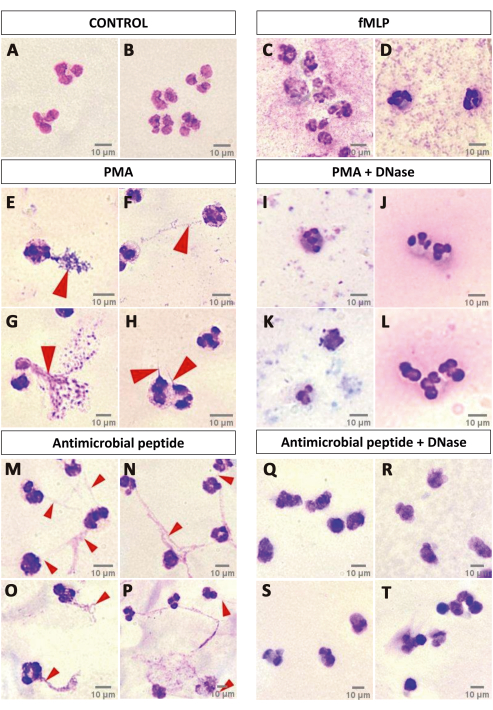
Figure 6: NET formation suggestive assay. A qualitative analysis of panoptic-stained neutrophils after 1 h of activation over a glass slide may indicate NET release. CTRL (A,B) and fMLP (C,D) groups did not show any indication of NET release, while activation with PMA (E–H) induced the formation of NET-like structures that were degraded by DNase I treatment (I–L). Investigation of the effects of an antimicrobial peptide on the neutrophils showed that such a stimulus might be able to elicit NET release, since the slides presented the NET-like structures (M–P) that were degraded by DNase I (Q–T). Red arrows point to NET-like structures. Scale bars: 10 µm. Please click here to view a larger version of this figure.
