The typical output obtained using the newly developed procedure in the 4-day-old de-etiolated Arabidopsis seedlings of wild-type (WT), ecotype Columbia-0 (Col-0) is shown in Figure 3. Under control conditions (DMSO-supplemented MS media), the chlorophyll biosynthetic curve starts with an initial burst of the chlorophyll synthesis, in which the protochlorphyllide pool synthesized during the scotomorphogenic phase of the growth, is quickly converted to chlorophyll owing to the light-induced PORs7,8,9. The initial phase of fast chlorophyll accumulation takes approximately 10 min and is followed by a lag phase, during which the minima of the dark-synthesized protochlorophyllide are reached (approximately 30 min after irradiation; for the HPLC-measured protochlorophyllide curve, see7). During the lag phase, the chlorophyll biosynthetic genes are upregulated10, leading to light-induced production of protochlorophyllide. The newly synthesized protochlorophyllide is promptly converted to chlorophyll, detectable as exponential phase (in case of WT Col-0 starting at approximately 120 min after irradiation), finishing with another lag phase at approximately 4 h after the de-etiolated seedling irradiation (Figure 3). In the presence of 6-benzylaminopurine (BAP), the first significant differences are detectable during the exponential phase7, suggesting negative effect of BAP on the chlorophyll kinetics in later stages of chlorophyll biosynthesis (at about 2 h after the start of illumination with actinic light; Figure 3).
For comparison of different conditions and/or genotypes, normalization of the raw data is necessary. As no chlorophyll was detectable using HPLC under various conditions and/or different genotypes in etiolated seedlings, we performed the normalization (F/F0) to the T0 fluorescence levels (F0) measured for the corresponding treatment and/or genotype7. To demonstrate the importance of normalization, we present both raw data and the data normalized to the mean chlorophyll fluorescence value of control measured at T0 (F0; Figure 3A and Figure 3B, respectively).
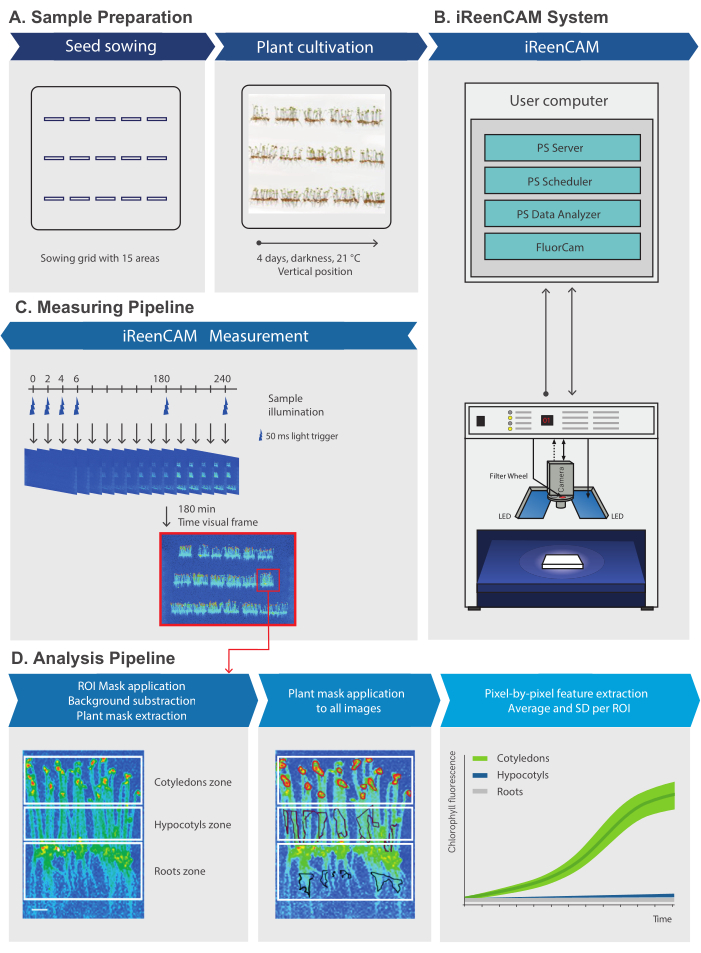
Figure 1: Measuring device protocol overview. The scheme of iReenCAM measurement and analysis pipeline. (A) Sample preparation by grid-defined seed sowing, stratification, light induction of germination and vertically oriented Petri plate cultivation in darkness. (B) Control module for automatic and programmable image acquisition and data management based on PlantScreen phenotyping SW toolbox organizes the operation of the entire system by controlling and synchronizing the HW operation with user defined measurement and analysis protocol. (C) The measuring protocol is designed for dynamic measurements of the fluorescent sample images in 2 min intervals for 4 h in total, i.e., 120 measuring rounds. Time visual frame of representative false-color image of vertically oriented 4-day-old Arabidopsis seedlings acquired at time 180 min is used for ROI mask generation. (D) Mask defining ROI for a tissue of interest (e.g., cotyledon, hypocotyl, or root zone) is applied on the time visual frame, background subtraction is performed and pixel by pixel fluorescence values for each ROI (defined by the plant mask) from all measurement rounds is extracted. Finally, the raw data (fluorescence F) is normalized to the mean fluorescence value at T0 (F0). Scale bars = 1 cm (A) and 0.25 cm (C). The figure was modified from7. Please click here to view a larger version of this figure.
Figure 2: Seed placement. The figure shows placing Arabidopsis seeds on the Petri plate with light-tight edges using the sawing grid. Please click here to view a larger version of this figure.
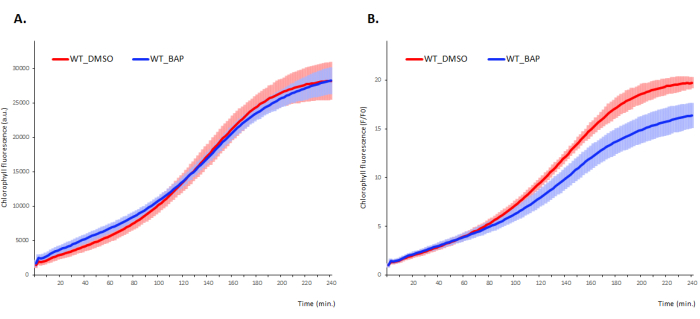
Figure 3: Chlorophyll accumulation kinetics in the early stages of Arabidopsis de-etiolation. Etiolated WT Col-0 seedlings were grown on media supplemented with BAP or DMSO (mock). (A) The mean value ± SD (shaded area), n=9 of raw data and (B) data normalized to the mean fluorescence value at T0 (F0). Please click here to view a larger version of this figure.
Supplementary Figure 1: Sowing grid. Left: Sowing grid with outlined rectangular boxes for placing the seeds of each genotype into a measuring spot located in the area of actinic light homogeneity (light intensity ≥ 0.7 of the maximum light intensity). Right: Schematic representation of 4 days old, etiolated Arabidopsis seedlings grown for the analysis. The sowing grid provides a possible scheme for Arabidopsis seeds positioning ensuring light homogeneity and proper seed density (each slot can be used for seed placement as the size of a slot, distance between the slots and the light homogeneity in the area of the grid are unified). Please click here to download this File.
Supplementary Figure 2: Fluorescence measuring protocol. Please click here to download this File.
Supplementary Figure 3: Experimental configuration allowing to avoid unwanted light exposure.(A) The measuring device is placed in the walk-in phytotron, (B) to the chamber separated by light-tight door. (C) The in vitro cultivation boxes (red arrowhead) dedicated for the etiolated seedlings growth under defined conditions (temperature and relative humidity) are placed just beneath the device (yellow arrowhead), ensuring the minimal risk of light exposure. The source of green dim light (blue arrowhead) is mounted on the wall next to the control PC (orange arrowhead). Please click here to download this File.
Supplementary Figure 4: Mask generation procedure in the workflow using PS data analyzer software. Print screenshots of individual steps to be performed for the tray mask (steps 4.3-4.6) and plant mask (step 4.12) generation and data analysis (steps 4.7 and 4.13). Please click here to download this File.
Supplementary Figure 5: Sowing density affecting measurement variability. Chlorophyll accumulation kinetics in 4-days-old, etiolated Arabidopsis WT Col-0 seedlings grown (A) separately (as individual seedlings, here n=5) or in a group of (B) high (HD, n=30-40) or (C) low density (LD, n=10-15). The n corresponds to the number of seedlings per slot of the sowing grid, data represent the mean values ± SD (shaded region). The high or low density corresponds to the number of seedlings per slot of the sowing grid as mentioned. Please click here to download this File.
Supplementary Figure 6: Cultivation interval and space requires species-specific optimization. Growth of various plant species using the Arabidopsis-optimized protocol. (A) Seeds placement on the sowing grid. (B) 4-day-old, etiolated seedlings of (from the left to the right) Arabidopsis thaliana, Brassica napus, and Crambe abyssinica. Please click here to download this File.
