1. Preparing Flame-Sealed Glass Micropipette
- Using microelectrode glass, pull a standard patch-clamp micropipette on a Flaming-Brown or equivalent micropipette puller (tip diameter ˜2 μm). Place micropipette tip into the flame from a Bunsen burner for ˜2 sec until a fused ball forms with a diameter of 200-300 μm.
- Place flame-sealed patch pipette onto holder on a micromanipulator that can be rapidly vibrated side-to-side (travel distance 100-200 μm) using a piezoelectric bimorph, relay, or equivalently effective device.
2. Preparing Brain Slices from P1-P21 Rats or Mice
- Prepare artificial cerebrospinal fluid (aCSF) with the following composition (in mM): 124 NaCl, 4.5 KCl, 1.2 NaH2PO4, 26 NaHCO3, and 10 D-glucose. Bubble solution with 95% oxygen/5% CO2 gas for ˜15 min, then add 2 mM CaCl2 and 1 mM MgCl2.
- Cutting brain slices:
- Anesthetize animal with halothane or isoflurane. Decapitate animal – remove brain – block brain in desired orientation (coronal, parasagittal, transverse, etc.) to include regions of interest.
- Affix the blocked brain to stage of a vibrating brain slicer submerged in aCSF – section brain at 250-400 μm thickness – place slices in aCSF bubbled with carbogen in a “preincubation” chamber on netting that allows for fluid/gas exposure on all surfaces – allow slices to equilibrate in this medium for at least one hour.
3. Vibrodissociation
- Fill a 35 mm diameter culture dish with HEPES-buffered saline solution of the following composition (mM): 150 NaCl, 5 KCl, 10 HEPES, 1 MgCl2, 2.5 CaCl2, 10 D-glucose with pH set to 7.4 using NaOH and osmolarity adjusted to ˜300 mOsM using sucrose. Culture dishes may be suspension dishes, standard cell culture dishes, culture dishes with glass coverslip inserts, or dishes coated with, e.g., poly-L-lysine, depending on the need for stronger cell adherence to the dish bottom.
- Place the slice in the culture dish and visualize with a dissecting stereoscope at 250 x. Hold slice to the bottom using a bent platinum wire (0.5 mm diameter) placed on the top surface of the slice to act as a weight.
- Position the tip of the flame-sealed micropipette on the slice surface in the desired brain region. Activate micromanipulator to vibrate the tip laterally at 10-30 Hz, with excursion distance ˜100 μm. Using the micromanipulator, move the tip deeper into the slice tissue such that it passes through the entire slice within ˜30 sec. Repeat this step as needed to maximize the number of isolated neurons obtainable from a given brain region.
- Remove the tip from the slice – pick up the slice with forceps and gently shake it while still in solution – then remove slice completely and discard.
- Allow the dissociated cells to settle and adhere to the dish bottom for at least 10 min.
4. Electrophysiological Recording
- Place the dish containing neurons on the stage of an inverted microscope and visualize with 10 x to 63 x objective. Look for neurons with a smooth patent membrane, no blebbing, and a detectable but not oversized nucleus. If phase contrast optics are used, look for phase-bright neurons with a yellowish-tinge, not too blue. Superfuse cells with extracellular solution containing desired salts, nutrients, receptor antagonists, etc. (our standard is the HEPES-buffered saline mentioned above (section 3.1)), often supplemented with 5 μM 2,3-dioxo-6-nitro-1,2,3,4-tetrahydrobenzo[f]quinoxaline-7-sulfonamide disodium (NBQX), and 25-100 μM D-2-amino-5-phosphonopentanoic acid (AP5) to block ionotropic glutamate receptors that allows for isolation of fast GABAA receptor-mediated IPSCs1,2.
- Pull a standard patch micropipette using a Flaming-Brown or equivalent puller. Pipette tip resistance should be 2-4 MΩ (depending on target cell size) when filled with a Cl– – based solution.
- Establish a whole-cell recording from a visualized neuron using standard electrophysiological techniques.
- Record spontaneous postsynaptic currents (sPSCs) using a gap-free data acquisition protocol. Apply 0.2-1 μM tetrodotoxin and/or low Ca2+ – containing extracellular solution to record miniature postsynaptic currents (mPSCs).
- Solutions and pharmacological agents can be directly applied to cells. We use local superfusion from fused square-tipped glass tubes with solution exchange involving lateral movement of the tubing mounted on a stepper-motor-driven micromanipulator. This allows for solution exchange in 10s to 100s of ms to achieve rapid changes in extracellular molecular content, application of receptor agonists, etc.
- Akaike and colleagues have developed techniques to stimulate single presynaptic boutons3,4. A micropipette is placed near a visualized bouton and stimulation is given (negative stimulus currents of 5-10 μA, 0.1-0.2 ms duration). Unitary IPSCs are recorded, and methods such as the method of failures can be used to examine changes in presynaptic and postsynaptic function.
5. Imaging Presynaptic Terminals
- Postsynaptic neurons can be filled with various dyes via the patch pipette. Neurons can also be made from mice expressing fluorescent proteins (FPs) such as green fluorescent protein (GFP) in selected cellular populations.
- Presynaptic terminals can be visualized on vibrodissociated neurons made from mice that express FPs in particular interneuron populations. For example, mice expressing GFP driven by the glutamate decarboxylase 65 (GAD65) promoter show green somata and processes in hippocampal basket cells and other interneurons. Vibrodissociated pyramidal neurons from the hippocampal CA1 region have GFP-positive axon terminals apposed to their somata and proximal dendrites (Figure 1A). Pyramidal neurons vibrodissociated from mice that express synaptopHlourin (a pH-sensitive GFP mutant fused to the synaptic vesicle-associated VAMP2 protein) under control of the Thy1 promoter also have FP-positive attached boutons that show pH-sensitive fluorescence (Figure 1B).
- Presynaptic boutons can be loaded with calcium indicator dyes and other fluorescent molecules using AM-esterified compounds5. Figure 2 shows an example of loading using the calcium-indicator dye Fluo4-AM. First, 1-2 μM AM-esterified dye is applied to neurons at 37° for 10 min. Dye can penetrate into the intracellular environment, where esterases cleave the molecule, yielding free, cell-impermeant and unquenched dye. This approach loads both pre- and postsynaptic cellular elements. Following the loading, the cells are washed with HEPES-buffered saline solution and kept at 37° for an additional 10 min. Prior to making a GΩ-seal with a glass pipette electrode, the cells are rinsed 3 times with external buffered solution.
- A whole-cell recording is then established using a dye-free intracellular solution. After 2-5 min of recording, the dye is mostly removed from the postsynaptic neuron, allowing visualization of Fluo-4-loaded synaptic boutons. This technique was pioneered by Ye et al5.
- The dye-loaded boutons can then be visualized using a high magnification, high numerical aperture objective with either a camera-based or multiphoton microscope. Simultaneous whole-cell recording and calcium imaging can be achieved using a setup as shown in Figure 3. The images of the neuron and boutons shown in Figure 2C were captured with an electron multiplying charge coupled device (EMCCD) camera. The power of the light source for excitation was attenuated to 1.2 % with a neutral density 1.0 filter and an iris filter adjusted to 12% output. Calcium transients from the green presynaptic boutons can be observed and recorded in real time, and measured offline.
6. Representative Results:
Spontaneous and Current Injection-Evoked Firing of Vibrodissociated Neurons
Recordings from a typical vibrodissociated hippocampal CA1 pyramidal neuron are shown in Figure 4A. Spontaneous overshooting action potentials can be observed in pyramidal neurons dissociated from rat basolateral amygdala, hippocampus and VTA1,5. During current-clamp recordings from CA1 pyramidal neurons, hyperpolarizing current injection produces typical voltage responses while depolarizing currents of sufficient magnitude elicit overshooting action potentials with the typical delayed firing pattern with moderate current levels and non-accommodating action potentials in response to stronger current injection (Figure 4B).
Spontaneous and Miniature GABAergic Inhibitory Postsynaptic Currents in Vibrodissociated Neurons
During voltage-clamp recordings with a CsCl-based intracellular solution, spontaneous synaptic currents are observed (Figure 5A). These events are eliminated in low calcium-containing solution2,6, and in most neuronal types are completely blocked by GABAA receptor antagonists such as bicuculline or gabazine (Figure 5B, C), indicating that these are sIPSCs mediated by GABA release and activation of this ionotropic receptor subtype. However, in principal neurons from brain regions such as the basolateral amygdala2 and the ventral tegmental area5,7, GABAA receptor blockade reveals shorter duration EPSCs mediated by glutamatergic activation of AMPA-type receptors. Interestingly, application of TTX at concentrations that are specific for blockade of the most toxin-sensitive voltage-gated sodium channels reduces the frequency and amplitude of the sIPSCs observed in vibrodissociated neurons from several brain regions (Figure 6A)2,4,7. Thus, sodium channel activity participates in GABA release in the pinched-off synaptic boutons. It is not yet clear if full-blown sodium action potentials occur in these terminals. It is worth noting that the input resistance of a well-sealed 1 μM diameter axon terminal is likely to be well into the GΩ range. Thus, opening of even a small number of sodium channels might be sufficient to depolarize terminals to activate the calcium channels that mediate excitation secretion coupling. On the subject of these calcium channels, evidence suggests that N and P/Q-type channels participate in release at GABAergic terminals in the vibrodissociated preparations from hippocampal CA1 and elsewhere8.
Modulation and plasticity of transmission by a number of neurotransmitters and receptors has been examined using vibrodissociated neurons3,4,9. The neurotransmitters shown to have such modulatory actions are adenosine, GABA, serotonin, endocannabinoids, etc2,6,10,11,12. In addition, short-term synaptic plasticity, such as endocannabinoid-dependent depolarization-induced suppression of inhibition (DSI) has also been described in this preparation2,11. These findings indicate that many forms of receptor-mediated and trans-synaptic signaling by endogenous neuromodulators are intact in the vibrodissociated preparation.
Calcium Transients and Vesicular Release in Axon Terminals in the Vibrodissociated Preparation
Using the EMCCD-equipped inverted microscope describe above, we have examined calcium transients in presynaptic terminals in the vibrodissociated neuron-bouton preparation. Figure 2 shows cell loading with AM-esterified Fluo-4 and subsequent postsynaptic dye dilution in a hippocampal CA1 pyramidal neuron. Spontaneous calcium transients are observed in some dye-filled boutons shown as regions of interest (ROIs) in Figure 6B. Application of 1 μM tetrodotoxin (TTX) eliminates these spontaneous transients, indicating that they are mediated by activation of voltage-gated sodium currents that are spontaneously activated in boutons, leading to subsequent calcium rises. Increasing extracellular KCl from 10 mM to 40 mM with fast solution exchange produces increased fluorescence as measured in ROIs that correspond to presynaptic terminals (Figure 7A). Simultaneous recording from the postsynaptic neuron is used to monitor sIPSCs and determine if event frequency is increased by high-K+ depolarization (Figure 7B).
To visualize vesicle fusion in the presynaptic boutons we have used mice expressing the synaptopHlourin construct under the control of the Thy1 promoter (labeled spH21 mice). SynaptopHluorin in a molecular construct in which ecliptic pHlourin (a GFP mutant with enhanced pH sensitivity)12 is linked to the vesicle associated membrane protein (VAMP2)13. This arrangement situates the pHlourin motif in the vesicle lumen where the relatively acidic environment quenches fluorescence. Upon vesicle fusion this motif is exposed to the more neutral extracellular environment with a resultant increase in fluorescence at puncta that correspond to vesicles/presynaptic terminals. Vibrodissociation of hippocampal neurons from spH21 mice allows for visualization of fluorescent puncta of the size and location expected for GABAergic terminals (Figure 8A). Application of the high-K+ – containing external solution increases fluorescence in these terminals, and this effect is blocked in the presence of an external solution in which extracellular calcium is reduced to 0.2 mM (Figure 8B). Thus, the depolarization-induced increase in fluorescence appears to reflect excitation-secretion coupling at GABAergic synapses in the neuron-bouton preparation.
The ability to measure presynaptic calcium transients and vesicle fusion in axon terminals of the neuron-bouton preparation allows us to examine effects of neuromodulators, drugs of abuse and synaptic plasticity on presynaptic mechanisms involved in excitation-secretion coupling and exocytosis/endocytosis. These techniques can also be combined with other molecular tools and genetically-engineered mice to examine the roles of particular proteins in presynaptic function and modulation/plasticity.
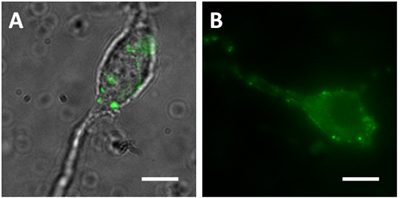
Figure 1. Vibrodissociated neurons from the hippocampal CA1 region (A) Merged image of DIC and green fluorescence images from a GAD65 mouse. DIC image is merged to clearly show the locations of the green terminals (B) Fluorescence image from a synaptophluorin (spH21) mouse. Scale bar = 10 μm
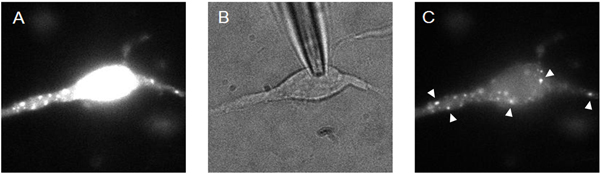
Figure 2. Calcium indicator-loading procedure: (A) whole cell is loaded with AM-esterified dye; (B) whole-cell recording dilutes dye; (C) terminals are visualized as regions of interest (arrowheads).
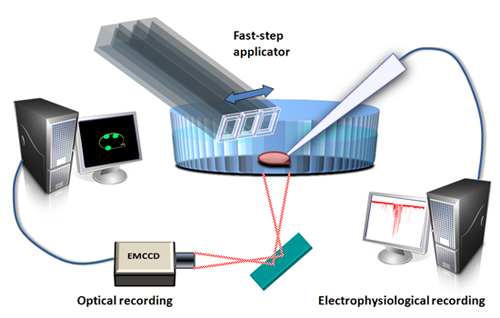
Figure 3. Schematic diagram of the experimental setup for simultaneous whole-cell recording and calcium imaging in presynaptic terminals on vibrodissociated neurons. EMCCD=electron multiplying charge coupled device.
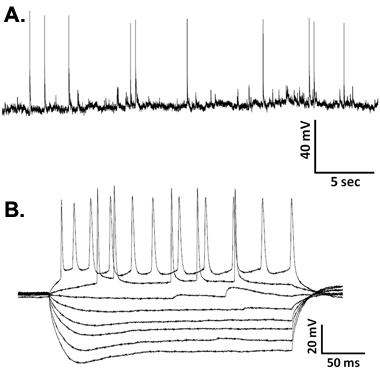
Figure 4. Representative waveforms showing (A) spontaneous action potentials from a vibrodissociated CA1 neuron and (B) membrane voltage responses to hyperpolarizing and depolarizing current injections in current-clamp recordings from a CA1 neuron.
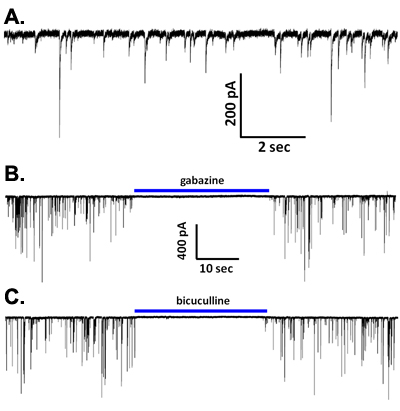
Figure 5. (A) Representative waveforms of spontaneous IPSCs from a CA1 pyramidal neuron. (B-C) The IPSCs were blocked by either gabazine (10 μM; B) or bicuculline (20 μM; C).
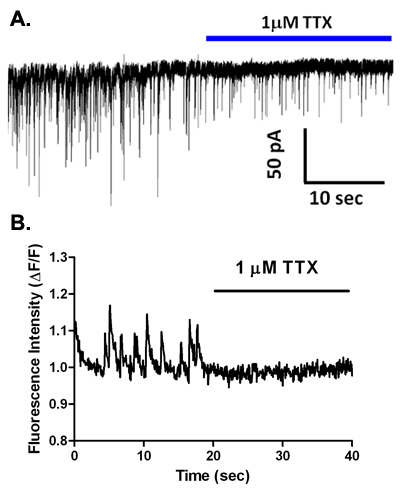
Figure 6. TTX inhibits spontaneous GABAergic synaptic transmission, and eliminates presynaptic calcium transients in the vibrodissociated hippocampal preparation. (A) Recording from a vibrodissociated neuron before and during TTX application. Note the decrease in frequency and amplitude of sIPSCs. (B) Calcium transients observed in a Fluo-4-loaded presynaptic terminal on a vibrodissociated neuron before and during TTX application. Note the complete loss of transients.
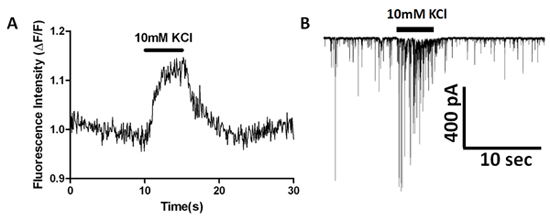
Figure 7. Simultaneous calcium indicator imaging and sIPSC whole-cell recording showing effects of high K+ stimulation. (A) Fluorescence measured over time from a presynaptic bouton loaded with Fluo-4AM dye. (B) Simultaneous increase of sIPSC frequency and amplitude during high K+ application.
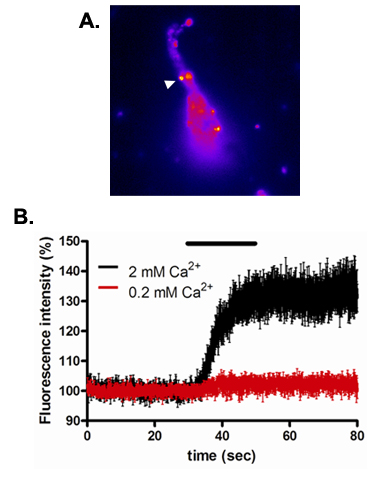
Figure 8. Ca2+-dependent high-K+ response in presynaptic boutons of spH21 pyramidal neuron from the hippocampal CA1 region. (A) Fluorescence image of a vibrodissociated neuron. (B) High-K+ application (indicated by black bar) resulted in a sustained increase in fluorescence in the presence of our normal extracellular Ca2+-containing solution (2 mM Ca2+). No fluorescence increase was observed at low extracellular Ca2+ (0.2 mM Ca2+). Data in graph were normalized to pre-high-K+ fluorescence levels, and show average response from 3 boutons.
