Odor-evoked responses of a single ORN to two different alcohols are shown in the Figure 3D. Depending on the recording location (sensilla type, placement of the electrode) multi-unit recordings can be achieved.
A raw extracellular waveform from an AL recording is shown in Figure 6A. Action potentials or spikes of varying amplitudes originating from different PNs can be observed in this voltage trace. Although the locust antennal lobe has excitatory projection neurons and inhibitory local neurons, only PNs generate sodium spikes that can be detected extracellularly3. This observation suggests that the multi-unit recording technique presented here can be used to selectively monitor the output of the antennal lobe circuits, thereby making locusts an attractive invertebrate model for studying olfactory coding.
An example of a mushroom body recording is shown in Figure 6B. Unlike the ORNs and PNs, the KCs have lower baseline activity and respond to odors in a sparse and selective manner.
To isolate single unit responses from these multi-unit recordings, we performed off-line spike sorting (with the best four channels) using published software implemented in IGOR Pro (Wavemetrics)12. Examples of PN and KC spike sorting are shown in Figure 6C, and D, respectively.
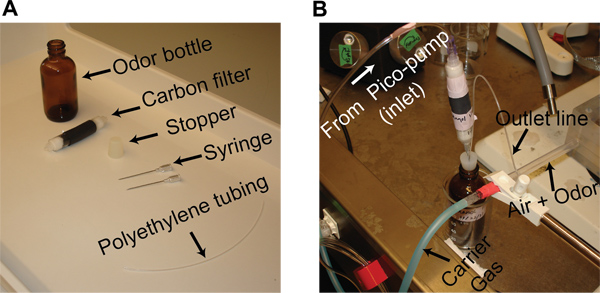
Figure 1. Odor stimulation. (A) All components needed for preparing an odor bottle are shown. (B) The inlet connection from the pico-pump and the outlet connection from the odor bottle to the odor delivery tube are shown. A constant stream of desiccated air is used as the carrier gas stream and is directed at the antenna during experiments.
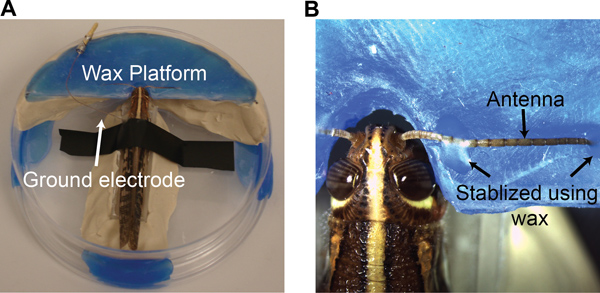
Figure 2. Preparation of a locust antenna for single sensillum recordings. (A) The locust is placed in a custom made chamber with a ground electrode placed in the gut. (B) A method to stabilize an antenna using a wax platform is shown.
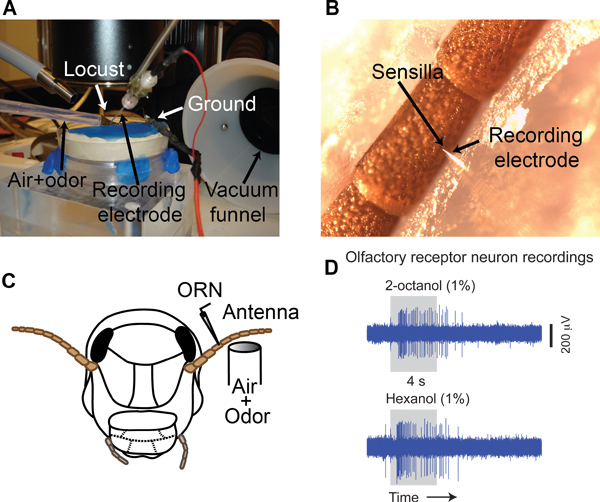
Figure 3. Single sensillum recordings. (A) A typical recording set up. A mixture of carrier gas and odor vapor is supplied through a delivery tube. ORN action potentials are recorded using a glass electrode. Delivered odorants are removed using a vacuum funnel situated right behind the antenna. (B) Electrode placement as seen through the stereomicroscope. Arrows indicate the placement of the glass electrode tip at the base of a sensillum. (C) A schematic of the single sensillum recording approach. (D) Raw extracellular voltage traces showing responses of an ORN to two different odors (2-octanol and 1-hexanol).
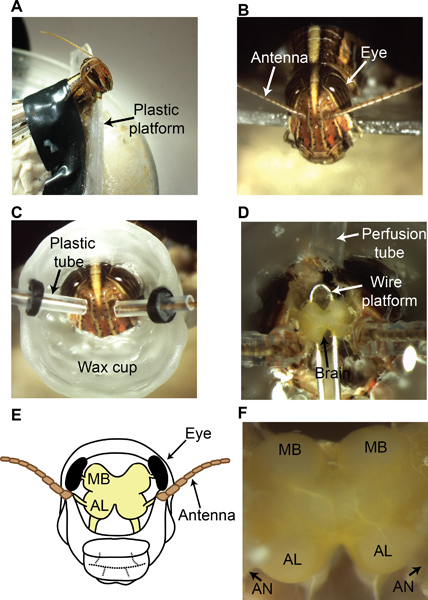
Figure 4. Locust dissection procedure. (A) A locust is restrained and positioned in a custom-designed dissection setup as shown. (B) View of the locust head from above. Both compound eyes and antennae can be clearly seen (C) A wax cup is built around the dissection site to allow saline perfusion during and after the dissection process. (D) An exposed locust brain is shown (the yellow-pigmented neural tissue). A platform is placed beneath the brain as shown to stabilize the brain. A saline perfusion tube is attached to the wax cup. (E) A schematic of the locust brain. (F) A magnified image of the locust brain after the dissection clearly showing the regions of interest: antennal lobes (AL) and the mushroom bodies (MB). The antennal nerve (AN) contains axon bundles that transmit the ORN action potentials from the antenna to the antennal lobe.
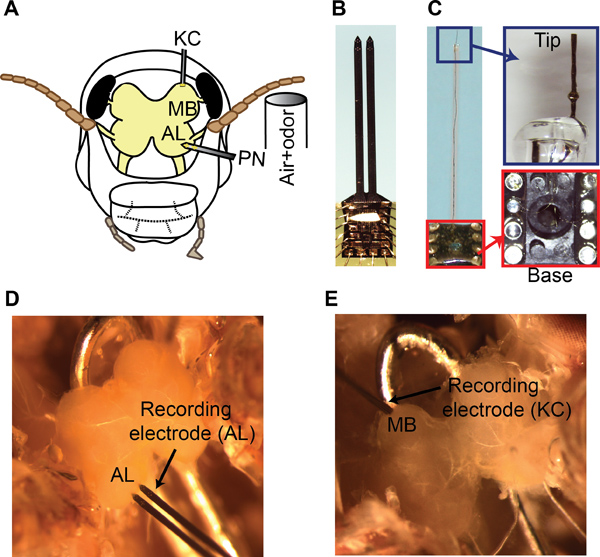
Figure 5. Multi-unit recordings from the antennal lobe and the mushroom body. (A) A schematic showing the recording configuration and the odor delivery setup. (B) A 16-channel NeuroNexus recording electrode used for PN recordings is shown. (C) Left panel, a custom made 8-channel twisted wire electrode is shown. Right panel, the electrode tip and the connections of wires to the IC socket are shown. (D) Placement of the 16-channel recording electrode in the AL. Only the bottom four electrodes in each shank are inserted into the tissue. (F) Placement of the twisted wire electrode in the superficial MB layers for KC recordings is shown.
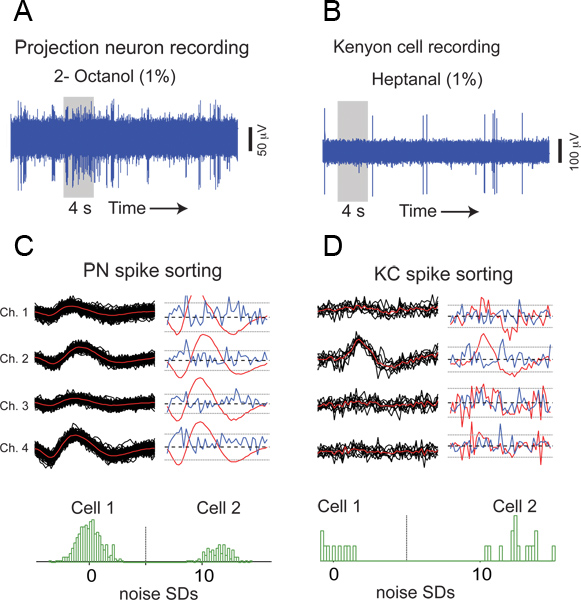
Figure 6. Representative results from an antennal lobe (AL) and a mushroom body (MB) recording. (A) A raw extracellular trace from a multi-unit AL recording is shown. A 4 s odor pulse was applied during the time period indicated by the gray box. (B) Similar plot but showing raw KC responses to an odor. (C) An example of PN spike sorting. Extracellular waveforms from four independent channels of a multi-channel electrode are shown for all spiking events arising from a single PN. Individual events (black), mean (red), and SDs (blue) are shown for both cells. Histograms obtained by projecting high-dimensional PN event representations (180 dimensional vector obtained by concatenating 3 ms signals from all four electrodes) onto the line connecting their means. To be considered a well-isolated unit, as in this case, a bimodal distribution with cluster centers at least five times the noise SD apart is expected for every pair of simultaneously recorded cells12. (D) An example of KC spike sorting is shown.
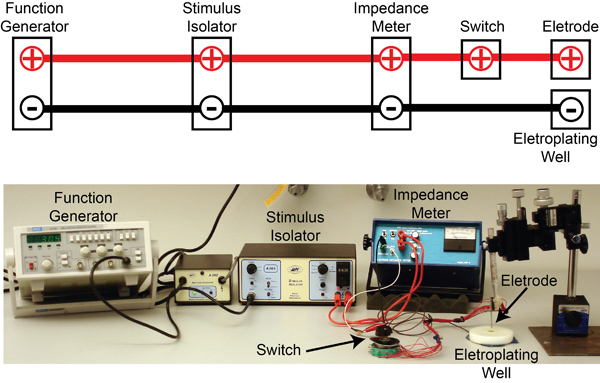
Figure 7. The electroplating set up: The circuit diagram showing connections between the different components are shown above a picture of the actual setup. Briefly, 3 Hz square pulses (5V amplitude) from a function generator (MCP, SG 1639A) are used to gate a stimulus isolator (WPI, A365) that then delivers 5 μA of current to an electrode impedance tester (BAK Electronics, IMP-2). The impedance tester can be operated to either test the electrode impedance or allow current pulses from the stimulus isolator to be applied to the electrode for gold plating. In both cases, the multi-unit electrode is kept immersed in an electroplating well containing gold solution. A switch allows selection of the electrode channel to be gold plated.
