Generation of Human Cardiomyocytes: A Differentiation Protocol from Feeder-free Human Induced Pluripotent Stem Cells
Summary
Pluripotent stem cells, either embryonic or induced pluripotent stem (iPS) cells, constitute a valuable source of human differentiated cells, including cardiomyocytes. Here, we will focus on cardiac induction of iPS cells, showing how to use them to obtain functional human cardiomyocytes through an embryoid bodies-based protocol.
Abstract
In order to investigate the events driving heart development and to determine the molecular mechanisms leading to myocardial diseases in humans, it is essential first to generate functional human cardiomyocytes (CMs). The use of these cells in drug discovery and toxicology studies would also be highly beneficial, allowing new pharmacological molecules for the treatment of cardiac disorders to be validated pre-clinically on cells of human origin. Of the possible sources of CMs, induced pluripotent stem (iPS) cells are among the most promising, as they can be derived directly from readily accessible patient tissue and possess an intrinsic capacity to give rise to all cell types of the body 1. Several methods have been proposed for differentiating iPS cells into CMs, ranging from the classical embryoid bodies (EBs) aggregation approach to chemically defined protocols 2,3. In this article we propose an EBs-based protocol and show how this method can be employed to efficiently generate functional CM-like cells from feeder-free iPS cells.
Introduction
Historically, the investigation of the genetic and molecular mechanisms driving human development and disease has been based on the generation of genetically modified animal models. However, numerous human phenotypes fail to be successfully replicated in mice, mainly because of the biological differences existing between the two species. On the other hand, access to human tissues may be limited and often do not allow enough material to be obtained for in-depth experimental studies. The field of cardiovascular biology suffers from both these limitations: the physiology of the human heart is significantly different from that of the mouse, and a significant quantity of heart tissue is accessible only post-mortem or during heart surgery. Finding an optimal source for the differentiation of functional human CMs has therefore become a central topic in cardiovascular biology, and much effort has been made to address this issue. Various cells types have been proposed, including skeletal myoblasts, local cardiac stem cells, bone marrow mononuclear cells, endothelial progenitors, and mesenchymal stem cells. However, data obtained using these cells have not been consistent 4.
The derivation of human embryonic stem cells (ESC) first 5 and the groundbreaking discovery of induced pluripotent stem (iPS) cells by Yamanaka and Thomson 6,7 later seemed to provide solutions: these cells have the ability to grow indefinitely and the potential to give rise to all cell derivatives of the three germ layers, including the CMs. The use of iPS cells offers further advantages: being derived from autologous adult cells, they carry the same genome as the individual or patient from which they are derived. They therefore allow the development of in vitro models that facilitate the investigation of human genetic diseases and mechanisms of development. In virtue of this characteristic, iPS cells can also overcome problems related to immune rejection and ethical issues 8. For these reasons, even though ESCs still represent the gold standard in the field of stem cell biology, researchers worldwide are now moving more toward iPS cell technology.
iPS cells are now being employed to model human disease in vitro for various conditions, including congenital cardiovascular disorders 9,10. Recently, application of iPS cell disease modeling has been performed for monogenic cardiac disorders (i.e. long QT syndromes, catecholaminergic polymorphic ventricular tachycardia) and pathologies in which cardiac defects are part of a complex phenotype (i.e. Leopard and Timothy syndromes, dilated cardiomyopathy). These reports confirmed that patient-specific iPS cells that are differentiated into CMs display similar phenotypic and functional characteristics as the disease in vivo.
However, there are still many challenges to improve the efficacy of inducing the iPS cells into the cardiac lineage. Spontaneous generation of CMs from human ESCs via aggregate formation called embryoid bodies (EBs) have proven successful 17. Since this discovery, many other methods have been proposed. Many groups have greatly enhanced the efficiency of the differentiation protocol and have moved toward chemically defined and animal-product-free culture reagents (see Mummery, C., et al., Circulation Research (2012) for a comprehensive review of all existing methods 3).
Nevertheless, the classical method based on EB aggregation still represents the most commonly employed for performing functional studies and investigating disease mechanisms. Our proposed protocol is based on the aggregation of iPS cells into EBs and culture in the presence of serum and ascorbic acid, which has been shown to enhance the cardiac differentiation process and to positively impact the maturation of these cells 18,19. In this article we will go through this methodology in detail and will show how to use feeder-free iPS cell lines for generating patient-specific iPS-derived CMs.
Protocol
1. Feeder-free Maintenance and Passaging of Human iPS Cell Lines
- Prepare the Matrigel-coated dishes. Thaw one vial of human ESC-qualified Matrix on ice for one hour and dilute it in 25 ml DMEM-F12 medium. Add 1 ml to each 35 mm plate (or equivalent amounts per surface area if other dishes are used), keeping everything on ice and ensuring that all plates and tubes are pre-cooled. Cover plates with aluminum foil.
Note: Matrigel stock concentrations vary by the batch. Dilution instructions are indicated on the data sheet.
- Keep plates on ice overnight and remove from ice the next day. Plates can be kept in the fridge up to one week before using.
- Prepare the mTESR1 complete medium (or thaw a vial of Nutristem medium) and the H-ESM (human ESC medium) according to the table below.
H-ESM FINAL CONC FOR 500 ml Knockout Serum Replacement (KSR) 20% 100 ml GlutaMAX 100X 2 mM 5 ml MEM non-essential amino-acids 0.1 mM 5 ml Penicillin-streptomycin 100 U/ml – 0.1 mg/ml 5 ml 2-mercaptoethanol 0.1 mM 900 μl B27 Supplement – without VitaminA 1% 10 ml N2 Supplement 1% 5 ml Knockout DMEM – 370 ml
Stock medium can be stored at 4 °C for no more than 2 weeks. To prepare complete growth medium, basic FGF is added just before feeding at a final concentration of 20 ng/ml.
Note: Complete H-ESM conditioned on mouse embryonic fibroblasts may be used instead of mTESR1 or Nutristem for iPS cells.
- Place coated plates in an incubator at 37 °C for 30 min to 2 hr prior to passaging.
- Cells should be passaged approximately every 5 days, even if they have not reached confluence. Colonies should not be touching.
- Replace growth medium with 1 ml of dispase (1 mg/ml, dissolve from a 10 mg/ml stock in PBS).
- Remove differentiating areas with pulled glass Pasteur pipettes. Areas that should be removed include crater-like or cystic structures in the center of colonies, and any areas where cells have become separated from colonies and flattened and seem to be migrating outwards.
- Incubate at 37 °C under a culture hood until colony borders become brighter and begin to detach (usually around 5-7 min). Discard dispase. Wash with 1 ml H-ESM and discard.
- In 1 ml H-ESM, use a cell lifter (spatula-like scraper) to harvest colonies and collect in a 15 ml Eppendorf tube. Rinse with 1 ml H-ESM and add to the tube. Spin at 1,000 rcf for 4 min and remove the supernatant.
- Resuspend pellet in 1 ml pre-warmed growth medium (either mTESR1 or Nutristem). Avoid breaking up clumps too much – pipette up and down less than 6-8x. Determine how many cells should be plated and remove unnecessary cells (usually 1:4 or 1:5 dilution), e.g. for 2 plates at 1:5, remove 600 μl, then add 1.6 ml fresh medium.
- Remove Matrigel from plates. Place cells drop by drop, distributing evenly on the plate. Avoid shaking the tube excessively, as this will cause cells to move towards the center.
- Wash tube with 1 ml medium and divide between plates. Final volume in each plate should be 1.5 -2 ml.
- Change medium everyday, except for the day after passaging. Although it is ideal to change the medium everyday, this may be occasionally changed at a frequency of once every two days if no more than 2-3 days have passed since the last passage without affecting pluripotency or differentiation potential.
- If the cell line must be frozen, re-suspend instead in 1-2 ml mFreSR freezing medium using a 2 ml pipette and place 1 ml/cryovial. Place vials in a cryobox at -80 °C and transfer to liquid nitrogen in 3-4 days.
Note: Manual microdissection of iPS cells must be performed under a stereomicroscope. An in vitro fertilization (IVF) workstation (e.g. an IVF Workstation from KSystem) integrated with a stereomicroscope allows manipulation of the cells in sterile conditions while they are kept on a heated surface area. If this is not available, a stereomicroscope can be moved under a regular laminar flow hood.
2. Embryoid Bodies Aggregation and Cardiac Induction
- Prepare EBM-20 medium according to the table below.
EBM-20 FINAL CONC FOR 500 ml Fetal Bovine Serum (origin South America) 20% 100 ml MEM non-essential amino acids 0.1 mM 5 ml Penicillin – streptomycin 100U/ml – 0.1 mg/ml 5 ml GlutaMAX 100X 0.1 mM 5 ml 2-mercaptoethanol 50uM 450 μl DMEM/F12 – 385 ml
Use of FBS of South America origin is critical for determining the differentiation efficiency: employment of other types of sera may affect the differentiation process. To prepare complete EBM-20, add ascorbic acid (AA) to a final concentration of 50 μg/ml. Complete growth medium can be stored at 4 °C for no more than one week. - Harvest iPS colonies as described above (steps 1.6 to 1.9).
- Resuspend gently in 1 ml EBM-20 with a 2 ml pipette. Avoid breaking up clumps too much – pipette up and down no more than 2-3x, checking the size of the colonies under the stereomicroscope. Plate on ultra-low attachment plates in 1:1 ratio (e.g. for a 35 mm dish, use 1 well of a 6-well plate). Rinse tube with 1 ml EBM-20 and add to plate.
Note: The size of the colonies affect the differentiation process, controlling efficiency and timing of cardiac induction. Aggregation of homogeneously-sized EBs have been shown to reduce the variability observed during differentiation.
- On day 3, change EBM-20 once using a microscope to avoid removal of EBs. Keep pipette close to edge of plate, and remove medium.
- On day 7, switch EBs to plates coated with 0.1% gelatin and previously placed at 37 °C for 30 min. If necessary, gelatin can be removed and plates left under a cell culture hood O/N. Add 35-40 EBs per 35 mm dish.
- Check EBs for beating areas and change EBM-20 twice per week. Mark out where beating areas are located on the top of the dish cover, to facilitate they localization under the stereomicroscope. Contracting areas should appear after approximately 10-20 days.
- When areas with spontaneous contraction appear, switch medium to EBM-2 (prepare as EBM-20, with 2% FBS instead) and isolate them as described below in section 3.
- Continue to check other areas for beating and change medium twice per week until beating areas are needed for further experiments.
Note: Efficiency of the differentiation process is highly dependent on several factors, the most critical of which are the specific cell lines and the batch of serum used to induce cardiac differentiation. To obtain the highest efficiency, test cell lines for cardiomyogenic potential and various batches of serum.
3. Beating Areas Isolation and Culture
- Coat 12-well plates (4 cm2 area) or the plates of interest with 5 μg/cm2 fibronectin, diluted in PBS to make enough volume to cover the entire surface completely (300 μl total per well in 12-well plates). Keep uncovered in cell culture hood and allow to dry. Plates can be wrapped and stored at 4 °C O/N.
- Remove beating area using a glass pulled Pasteur pipette and scalpel, as needed. Transfer to fibronectin-coated plates with a 1 ml pipette.
- Add 1 ml EBM-2.
- Cross off the plate to indicate the areas where cells were removed and change the medium. Plates can be kept for up to 1 month, as other areas may start beating later on.
Note: If smaller beating clumps are needed, pipette the explanted area up and down several times under the stereomicroscope, until it breaks into smaller pieces. This will result in clusters of a few beating cells (see movie 3).
- Change medium twice a week until ready for the analyses.
Note: Cardiomyocytes obtained from this protocol possess fetal-like morphological and functional characteristics; maturation towards an adult-like phenotype may be obtained by increasing the time in culture to further differentiate these cells.
4. Single Beating Cells Isolation and Analysis
- Coat 2-chamber slides (4 cm2 area) or other appropriate supports with 5 μg/cm2 fibronectin diluted in sufficient PBS to cover the entire culture surface. If a glass support is used, coat with laminin and fibronectin ( 5 μg/cm2 each).
- Remove beating area with a Pasteur pipette/scalpel from the plates from section 3.
- Using a 1 ml pipette, transfer cells to a 15 ml tube containing 500 μl collagenase II (480 U/ml in PBS with Mg and Ca) and incubate 15 min at 37 °C, shaking the tube once during the incubation.
- Pipette up and down with a 200 μl pipette. If any clumps remain, transfer to fresh collagenase II and repeat step 4.3.
- Repeat step 4.4 until no large clumps are left.
- Inactivate collagenase with 3 ml EBM-2 (no AA). The tube can then be left under the hood in a heated rack until other tubes are ready. Centrifuge at 1,000 rcf for 4 min.
- Resuspend pellet in 1 ml 0.25% trypsin/EDTA (diluted from a 0.5% stock in 1X PBS) and incubate at 37 °C for 5 min. Combine fractions of digestions from the same initial sample.
- Inactivate trypsin with 4 ml EBM-2 (no AA). Pass cells through an 18G needle on a 2.5 ml syringe no more than 7x. Centrifuge at 1,000 rcf for 4 min.
- Resuspend cell pellet in 1 ml EBM-2 per well and plate. Cells can be used for functional and morphological analyses 2-3 days after plating.
Representative Results
In our studies we generated CMs from several iPS cell lines. These lines were initially generated on an MEF feeder layer but were immediately placed on Matrigel using a defined medium (either mTESR1 or Nutristem). Various assays were used to verify maintenance of the morphological properties, the expression of markers typical of pluripotent cells (OCT-4, SSEA4 and TRA1-60) and their genome stability (Figure 1).
Induction into the cardiac lineage was triggered by aggregation of iPS cells into EBs and culture in the presence of a specific type of serum and AA (Figures 2A and 2B). Contracting areas represented 20-35% of the total, depending on the cell line used (Figure 2C and Movie 1). Contracting CMs isolated from these regions by enzymatic digestion (Movie 2) expressed typical cardiomyocyte structural proteins (cardiac troponin, α-sarcomeric actin, myosin heavy chains α and β), gap junction proteins (connexin-43), and the major ion channels (Figures 2D-2F). Electrophysiological analysis of these cells revealed a heterogeneous population comprising cells with nodal, atrial, and ventricular profiles (data not shown, Priori, S.G., et al., J. Clin. Invest., In Press, 14,15,20).
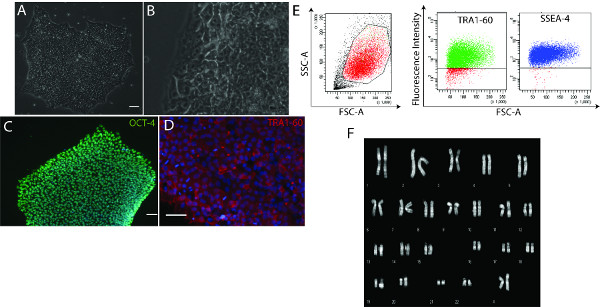
Figure 1. Feeder-free iPS cells maintenance does not affect pluripotency. A-B) iPS colonies grown on Matrigel retain their human ESC-like morphology, with phase-bright edges and prominent nucleoli. C-D) Immunofluorescence analyses showing iPS cells correctly expressing the transcription factor OCT-4 (C) and the TRA1-60 surface antigen (scale bars: 50 μm) (D). (E) Cytofluorimetric analysis confirmed the expression of typical surface pluripotency antigens (SSEA-4 and TRA1-60). The gating strategy is shown by the left scatter. (F) Representative karyotype analysis of iPS cells grown on Matrigel. Click here to view larger figure.
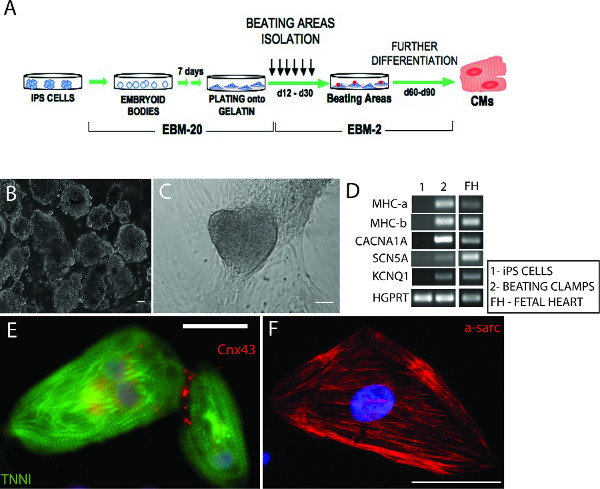
Figure 2. Differentiation of functional iPS-derived CMs. A) Schematic representation of the differentiation protocol. B) Embryoid bodies aggregated from iPS cells. C) Representation of an area of contraction. D) RT-PCR showing the expression of some cardiac-specific ion channels by the iPS-derived beating clumps (CACNA1A – calcium channel, voltage dependent, alpha 1A subunit; SCN5A – sodium channel, voltage-gated, type V, alpha subunit; KCNQ1 – potassium voltage-gated channel, KQT like subfamily, member 1) and both forms of myosin heavy chain (MHC-a and MHC-b). No expression was detectable in iPS cells, whereas cDNA from fetal heart cells was used as a positive control. HGPRT was used as housekeeping gene. E-F) Immunofluorescence staining showing single iPS-derived CMs expressing the structural protein α-sarcomeric actin, cardiac troponin I (TNNI), and the cardiac-specific junction connexin 43 (Cnx-43). Scale bars: 50 μm. Images were acquired using an Axiovision Zeiss fluorescence inverted microscope and analysed with ImageJ. Click here to view larger figure.
Movie 1. Contracting area derived from iPS cells. Click here to view movie.
Movie 2. Single contracting iPS-derived CMs, obtained from enzymatic digestion of a contracting area. Click here to view movie.
Movie 3. Small contracting cluster, obtained by partial mechanical dissection. Click here to view movie.
Discussion
Pluripotent stem cells have the potential to differentiate spontaneously into CMs, albeit with low efficiency and high variability among lines. The development of novel induction methods has therefore focused on improving the efficiency of the process and moving toward more defined protocols. However, most of these new differentiation methods are quite complex, requiring elaborate culture conditions, fine control of timing and concentration of reagents, and treatment with expensive cytokines and growth factors. Also, differences in the endogenous cytokine signaling levels of various iPS cell lines render it difficult to standardize cytokine concentrations, which often must be adjusted to appropriate levels for each cell line.
Maturation of the obtained iPS-derived CMs also represents a critical aspect to consider when attempting cardiac differentiation of iPS cells for disease modeling and drug discovery applications. The differentiation of human ESCs and iPS cells usually gives rise to CMs with a fetal phenotype i.e. without a completely mature structure. This is particularly true when monolayer approaches are used, and as a result these are more suitable for generating, amplifying, and isolating specific cardiac progenitor populations 2,3.
In this scenario, the “classical” EBs aggregation method, which is still widely used for disease modeling purposes, seems to better fit the experimental needs. This method is straightforward, economical, easily practicable, and suitable for all lines. There is no need for additional growth factors or cytokines, or for single-cell digestion, which are often not very well tolerated by iPS cells; also, they require additional treatments with ROCK inhibitors 21. Furthermore, the aggregation into EBS resembles the natural developmental process of a pluripotent cell, with the EBs providing a 3-dimensional environment and paracrine signals that are more similar to physiological conditions. Moreover, the addition of AA has been shown consistently to enhance cardiac differentiation of iPS cells, and also to improve their structural and functional maturation 18,19. Choosing iPS cell lines based on their cardiogenic potential, together with the selection of the most appropriate batch of serum, may further improve the efficiency of CM generation.
Finally, the method proposed here has also the advantage of not requiring mouse feeder cells, which eliminates the possibility of contamination with mouse cells and also further simplifies the technical procedures for iPS cell passaging and EBs formation. This method confers numerous advantages and constitutes a significant improvement over prior techniques, opening the way to future cardiovascular regenerative medicine applications.
Disclosures
The authors have nothing to disclose.
Acknowledgements
BS was supported by University of Milan-Bicocca Summer Student Research Training Program; research was supported from funds of the Italian Ministry of Health and Italian Ministry of Education, University and Research and Fondazione Humanitas to GC; we thanks Michael V.G. Latronico for critically reading the manuscript.
Materials
| Name of the reagent | Company | Catalogue number | Comments (optional) |
| Media and Reagents | |||
| Human ESC-qualified Matrix | BD Biosciences | 354277 | Thaw the 5 ml bottle at 4 °C on ice O/N, aliquot in pre-chilled cryo-tubes on ice, according to the data sheet dilution instructions and store at -20 °C . |
| Fibronectin, from human plasma, 0.1% solution | Sigma | F0896-2MG | Working concentration: 5 μg/cm2 |
| Laminin | Sigma | L2020-1MG | Working concentration: 5 μg/cm2 |
| PBS (no Mg and Ca), 10X | Lonza | BE17-515F | |
| PBS (with Mg and Ca), 10X | Bio Sera | XC-S2067/500 | |
| Dispase II, neutral protease, grade II | Roche | 4942078001 | Stock: 10 mg/ml in DMEM-F12 at -20 °C, working solution: 1 mg/ml in PBS (no Mg/Ca) |
| Trypsin/EDTA, 0.5% | Sigma | T3924 | |
| Collagenase type 2 | Worthington | 4176 | Dilute to 480 U/ml (e.g. 1.6 mg/ml) in PBS with Mg and Ca |
| DMEM-F12 | Life Technologies | 21331-020 | |
| Knockout DMEM | Life Technologies | 10829-018 | |
| Knockout Serum Replacement (KSR) | Life Technologies | 10828-028 | Thaw at 4 °C, aliquot and store at -20 °C |
| Foetal Bovine Serum (origin South America) | Invitrogen | 10270-106 | Batches should be tested for their cardiogenic potential. Thaw at 4 °C, heat-inactivate at 56 °C for 30 min, aliquot and store at -20 °C |
| PenStrep, penicillin-Streptomycin, 100X | Life Technologies | 15140-122 | |
| GlutaMAX, 100X | Life Technologies | 35050-038 | |
| MEM NEAA, 100X | Invitrogen | 11140-135 | |
| N2 supplement, 100X | Life Technologies | 17502-048 | |
| B27 supplement, 50X | Life Technologies | 12587-010 | |
| 2-mercaptoethanol | Invitrogen | 31350-010 | |
| Gelatin, from porcine skin | Sigma | G1890-100G | Make stock at 1% in water, store at -20 °C. Dilute to 0.1% in PBS and keep at 4 °C |
| mTeSR1 | Stem Cell Technologies | 5850 | Combine Supplement 5X with the basic medium, aliquot and store at -20 °C. Do not keep complete medium at 4 °C for more than 1 week |
| Nutristem hESC XF | Stemgent from Biological Industries | 05-100-1A | Thaw at 4 °C O/N, aliquot and store at -20 °C until use. Aliquots may be kept at 4 °C for no more than a week. |
| mFreSR | Stem Cell Technologies | 5853 | |
| Ascorbic acid | Sigma | A4544-25G | Prepare stocks at 10 mg/ml in water. Store aliquots at -20 °C in the dark. Add to EBM at the final concentration (500X) |
| Plasticware | |||
| 35 mm culture dishes | Becton Dickinson | 353001 | |
| Cell scraper | Corning Incorporated | 3008 | |
| 12-well plates | Becton Dickinson | 353043 | |
| Permanox 2-well chamber slides | Thermo Fisher Scientific | 177437 | |
| Glass 2-well chamber slides | Thermo Fisher Scientific | 177380 | |
| 6-well plates, ultra-low-attachment surface | Corning Incorporated | 3471 | |
| 18G needle | Becton Dickinson | 304622 | |
| Glass pasteur pipettes, 230 mm | VWR International | 612-1702 | |
| 2.5 ml syringe | Becton Dickinson | 300188 | |
| Equipments | |||
| IVF Workstation | KSystem, by Nikon | ||
| Nikon SMZ1500 Stereomicroscope | Nikon |
References
- Di Pasquale, E., Latronico, M. V., Jotti, G. S., Condorelli, G. Lentiviral vectors and cardiovascular diseases: a genetic tool for manipulating cardiomyocyte differentiation and function. Gene Therapy. 19, 642-648 (2012).
- Laflamme, M. A., Murry, C. E. Heart regeneration. Nature. 473, 326-335 (2011).
- Mummery, C. L., et al. Differentiation of human embryonic stem cells and induced pluripotent stem cells to cardiomyocytes: a methods overview. Circulation Research. 111, 344-358 (2012).
- Wu, S. M., Chien, K. R., Mummery, C. Origins and fates of cardiovascular progenitor cells. Cell. 132, 537-543 (2008).
- Thomson, J. A., et al. Embryonic stem cell lines derived from human blastocysts. Science. 282, 1145-1147 (1998).
- Takahashi, K., et al. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell. 131, 861-872 (2007).
- Yu, J., et al. Induced pluripotent stem cell lines derived from human somatic cells. Science. 318, 1917-1920 (2007).
- Yamanaka, S. A fresh look at iPS cells. Cell. 137, 13-17 (2009).
- Josowitz, R., Carvajal-Vergara, X., Lemischka, I. R., Gelb, B. D. Induced pluripotent stem cell-derived cardiomyocytes as models for genetic cardiovascular disorders. Curr. Opin. Cardiol. 26, 223-229 (2011).
- Park, I. H., et al. Disease-specific induced pluripotent stem cells. Cell. 134, 877-886 (2008).
- Carvajal-Vergara, X., et al. Patient-specific induced pluripotent stem-cell-derived models of LEOPARD syndrome. Nature. 465, 808-812 (2012).
- Moretti, A., et al. Patient-specific induced pluripotent stem-cell models for long-QT syndrome. N. Engl. J. Med. 363, 1397-1409 (2010).
- Yazawa, M., et al. Using induced pluripotent stem cells to investigate cardiac phenotypes in Timothy syndrome. Nature. 471, 230-234 (2011).
- Itzhaki, I., et al. Modeling of catecholaminergic polymorphic ventricular tachycardia with patient-specific human-induced pluripotent stem cells. Journal of the American College of Cardiology. 60, 990-1000 (2012).
- Jung, C. B., et al. Dantrolene rescues arrhythmogenic RYR2 defect in a patient-specific stem cell model of catecholaminergic polymorphic ventricular tachycardia. EMBO Molecular Medicine. 4, 180-191 (2012).
- Sun, N., et al. Patient-specific induced pluripotent stem cells as a model for familial dilated cardiomyopathy. Science Translational Medicine. 4, 130ra147 (2012).
- Kehat, I., et al. Human embryonic stem cells can differentiate into myocytes with structural and functional properties of cardiomyocytes. The Journal of Clinical Investigation. 108, 407-414 (2001).
- Cao, N., et al. Ascorbic acid enhances the cardiac differentiation of induced pluripotent stem cells through promoting the proliferation of cardiac progenitor cells. Cell Research. 22, 219-236 (2012).
- Takahashi, T., et al. Ascorbic acid enhances differentiation of embryonic stem cells into cardiac myocytes. Circulation. 107, 1912-1916 (2003).
- Moretti, A., et al. Patient-specific induced pluripotent stem-cell models for long-QT syndrome. N. Engl. J. Med. 363, 1397-1409 (2011).
- Watanabe, K., et al. A ROCK inhibitor permits survival of dissociated human embryonic stem cells. Nature Biotechnology. 25, 681-686 (2007).
