Protocol sections 1 and 2 detail how test groups can be obtained to study attributes of accelerated, slowed and reversed aging in colonies with a single age cohort. To monitor worker-type differentiation that accompanies the normal ontogeny we assessed forager counts (“entrance counts”) for 6 colonies (Figure 1, compare section 1). The graphs show that considerable change from nurse to the forager state is typically not observed before individuals are more than 10 days old. Marked variability in forager counts was observed with regards to the timing of foraging onset among different colonies, and as a marked day-to-day variation within each colony. Apart from colony specific demographic factors, such as different brood load, much variability is explained by changing weather conditions (time points marked in red in Figure 1). Close monitoring of foraging dynamics therefore is advised to optimize marking and collection efforts during the experiment.
The reversed ontogeny (section 2) from foraging back to nursing tasks can be validated by inspecting brood combs that are introduced into the forager-derived colonies (see steps 2.2 and 2.3). For three replicates Figures 2A, C, and E show brood combs before introduction into forager-derived colonies. Figures 2B, D, and F show the respective combs after removal. Patches of newly capped brood, healthy larvae, and increased pollen storage around brood cells indicate that former foragers now had successfully performed typical nest, including nursing tasks.
Lipofuscin (section 3) is a highly conserved symptom of cellular senescence, and can be readily assessed for post-experimental analyses in the various bee tissues. Figure 3 contrasts lipofuscin accumulation, measured as granule size (Figure 3E), in the hypopharyngeal glands of age matched nurse and forager bees. The difference in chronological age between the two young and the two old groups was ≥17 days, with only one group (foragers) spending these ≥17 days with outside flight and food collection activities. Representative microscopic images (Figures 3A-D) show increased lipofuscin accumulation only for the group of older foragers after more than 17 days of foraging (Figure 3D), not for older nurse bees of similar chronological age (37-43 days; Figure 3B). A two-factorial ANOVA with the fixed main factors worker-type (foragers, nurses) and age difference (Δage ≥17 days) revealed significant effects for worker-type, age difference and the interaction between both factors (Ftype = 33.67, P<0.001; FΔage = 21.93, P<0.001; Ftype x Δage = 22.07, P<0.001). However, post-hoc tests showed significant effects only when contrasting older foragers (≥17 days of foraging) to younger foragers, or to both nurse groups (PF17d vs. F1d/N1d/N17d<0.001, Fisher’s LSD; Figure 3E). No difference was detected among the latter three groups, including chronologically young and old nurse groups (all tests with P >0.5, Fisher’s LSD; Figure 3E). This suggests that lipofuscin accumulation depends on forager specific activities (foraging age), rather than being function of chronological age only per se.
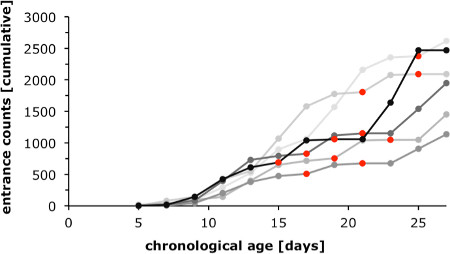
Figure 1. Worker-type differentiation during normal ontogeny. The graph displays entrance counts of foragers returning from foraging flights counted for 6 different colonies beginning 5 days after they were established (for details compare Protocol section 1.4). Considerable transition from nest to foraging activities was first observed when marked individuals of the single age cohort were about 10 days old. Varying slopes for the cumulative entrance counts indicate that the dynamics of the nurse bee to forager transition differ between colonies, and are affected by climatic factors. For example, on days with rain and less than two hours of foraging, the increase in entrance counts typically flattened out (data points in red).

Figure 2. Validating behavioral reversion. To test if foragers have successfully reverted to nursing tasks, we compared brood combs before they were introduced into forager-derived hives, and after they were removed from these hives. Representative images show brood combs before introduction (A, C, E) and after removal (B, D, F) from three different forager-derived hives, respectively. Brood care by previous forager bees is indicated by an increasing number of cells with capped brood (B, D, F; black arrow, inset in D), sustained survival of larvae in open cells (red arrow) and increased storage of pollen close to brood cells (white arrow). Note that forager-derived-colonies initially are typically less efficient in tending the brood than nurse-derived colonies. This can lead to higher larval mortality in the forager-derived-colonies. Pictures in B, D, F were taken 5, 4 and 7 days after brood combs were introduced into forager-derived colonies.
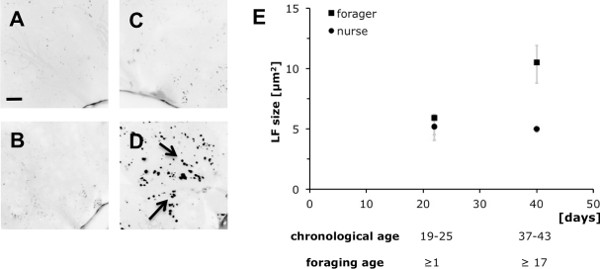
Figure 3. Accumulation of lipofuscin, a biomarker of cellular senescence, can indicate worker-type specific tissue deterioration. Representative microscopic images of hypopharyngeal glands in young (A) and old nurse bees (B), as well as in age-matched forager bees with ≥1 day (C), respectively ≥17 days foraging experience (scale bar in A = 20 μm). Lipofuscin accumulation was measured as granule size, and is given as medians and quartiles for N = 5 individuals for each age and worker-type (E). Foraging for 17 days resulted in significant lipofuscin accumulation, while the same period did not lead to lipofuscin changes in nurse bees (for statistics see Results).
