Membrane SPINE analysis allows the co-purification of membrane proteins and transiently interacting protein partners. The co-purification is achieved by using the cross-linking agent formaldehyde. Two parameters are critical to prevent unspecific cross-links: the formaldehyde concentration and the cross-linking time. The sufficient, but not excessive use of formaldehyde can easily be controlled by immunoblotting. Formaldehyde cross-linked protein complexes can be separated by boiling but not by SDS treatment. Hence, they can be visualized after blotting of the Strep-tagged membrane bait protein as smear in the upper section of the immune-blot of an unboiled sample.
A representative result of a Membrane SPINE assay, including all required controls, is presented in Figure 2. As bait protein the integral membrane protein CpxA of Escherichia coli was used6,11. CpxA is a sensor kinase and consists of an N-terminal sensor domain with two transmembrane domains (TMD) integrating a large extracytoplasmic sensor domain and a C-terminal highly conserved cytoplasmic catalytic domain12. After stimulation, CpxA activates its cognate response regulator CpxR. Activated CpxR diffuses off to mediate the response. For Membrane SPINE, the Strep-tag was fused to the C-terminus of CpxA (CpxA-Strep). CpxA-Strep cross-linked to other proteins or protein complexes is detected as a smear in the unboiled formaldehyde-treated sample (Figure 2, line 1 versus line 3) indicating sufficient cross-linking. Moreover, a direct protein-protein interaction of CpxA-Strep with its cognate response regulator protein CpxR as the prey protein is only detectable in the presence of formaldehyde (Figure 2, line 2 versus line 4) supporting the specificity of formaldehyde cross-linking.
A representative result of a Membrane SPINE analysis is presented in Figure 3. Samples corresponding to those of Figure 2 were silver stained. The arrow marks a band that is specific for the boiled sample. Due to the background, MS analysis also identified other proteins besides CpxR6. Therefore, the non-formaldehyde treated sample should always be analyzed, to assign background noise and specific interaction partner.
Table 1: Buffers and reagents required for membrane-SPINE.
| Buffer / Reagent / Medium | Working concentration | comment | |
|---|---|---|---|
| LB | 10 g Tryptone 5 g yeast extract 10 g NaCl to 1 L, adjust pH to 7.0 |
Luria Broth medium | |
| IPTG | 1 M Isopropyl-β-D-thiogalactopyranoside | 0.5 mM | |
| Tris-buffer | 20 mM Tris-HCl, pH 8 | Adjust pH to 8.0 using NaOH | |
| 0,1 M EDTA, pH 8 | 0.1 M EDTA | Adjust pH to 8.0 using NaOH | |
| PMSF | 1 M Phenylmethylsulfonylfluoride in 100% Isopropanol | 10 mM | PMSF is stable in 100% isopropanol but not in water! Stock solution can be stored at -20 °C; PMSF has to be adapted to room temperature before diluting in buffer; PMSF containing buffers should be used in within 10 min of preparation. |
| P1 | 20 mM Tris-HCl, pH 8.0 0.5 M Sucrose |
||
| P2 | 2 mg/ml lysozyme in 0.1 M EDTA, pH 7.5 | ||
| P3 | 20 mM Tris-HCl, pH 8.0 10 mM PMSF |
Use immediately after preparation | |
| Detergent | 20% Triton X-100 | 2% for solubilization | |
| Buffer W | Fill up 10 ml 5x concentrate to 50 ml add 150 µl 20% Triton X-100 |
100 mM Tris-HCl, pH 8.0 150 mM NaCl 1 mM EDTA 0.06% Triton X-100 |
5x concentrate is part of Strep-tag protein purification buffer set (IBA) |
| Buffer E | Fill up 1 ml 5x concentrate to 10 ml add 30 µl 20% Triton X-100 |
100 mM Tris-HCl, pH 8.0 150 mM NaCl 1 mM EDTA 2.5 mM Dethiobiotin 0.06% Triton X-100 |
5x concentrate is part of Strep-tag protein purification buffer set (IBA) |
| 5x SDS-PAGE loading dye | 0.3125 M Tris-HCl, pH 6.8 10% SDS 0.5 M DTT 50% glycerol 0.05% bromophenol blue |
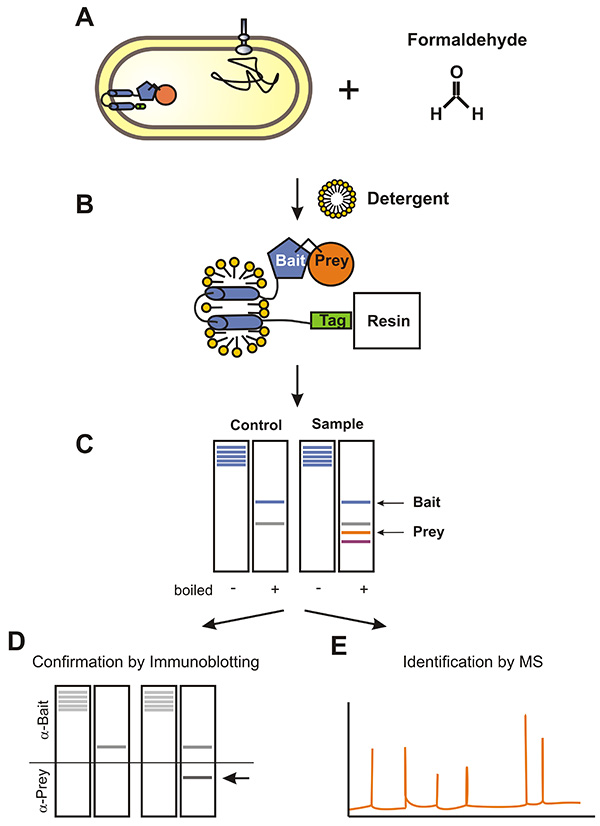
Figure 1. Flow chart of the Membrane-SPINE procedure using an Escherichia coli membrane protein as bait protein. A) Bacteria expressing the Strep-tagged membrane bait protein are treated with formaldehyde. Formaldehyde penetrates membranes and cross-links prey proteins to the membrane bait protein. B) The membrane fraction is prepared and membrane proteins are solubilized by detergent treatment. Subsequently, prey proteins are co-purified with the bait protein. C) Formaldehyde cross-links are reversed by boiling and proteins are separated by SDS-PAGE. Finally, prey proteins are either monitored by immunoblotting (D) or identified by MS-analysis (E). Click here to view larger image.
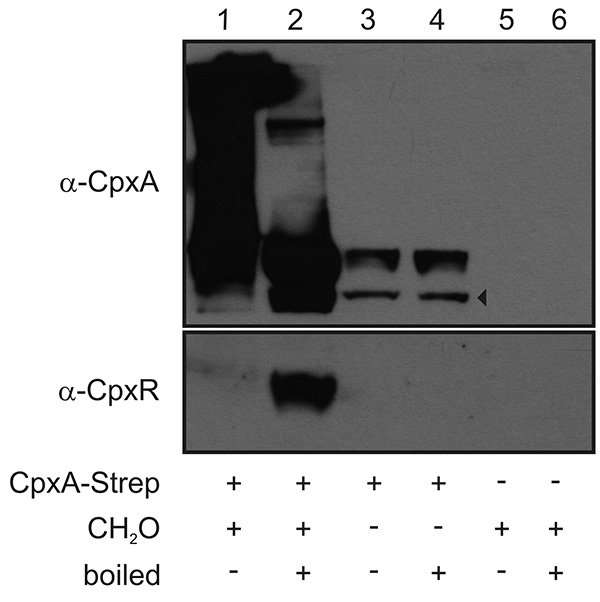
Figure 2. Representative immunoblot used to monitor a PPI of a membrane protein. Bacteria producing CpxA-Strep from a plasmid as membrane bait protein, were grown in LB medium to an OD600 of 1 and exposed to formaldehyde (CH2O) for 20 min. The inner membrane fraction was prepared, membrane proteins were solubilized by detergent treatment and CpxA-Strep was purified (lanes 1 and 2). Bacteria producing either CpxA-Strep without formaldehyde treatment (lanes 3 and 4) or carrying the empty vector with formaldehyde treatment (lanes 5 and 5) served as controls. Aliquots of each sample were boiled at 95 °C for 20 min (lanes 2, 4, and 6) to separate cross-linked proteins from CpxA-Strep. Proteins were separated in a 12.5% SDS-PAGE. Immunoblotting was performed and the blot was separated according to the size of CpxA-Strep (51 kDa) and the respective prey proteins CpxR (26 kDa). The two parts of the immunoblot were incubated with polyclonal antibodies raised against CpxA and CpxR, respectively. Subsequently, the blots were further treated with an anti-rabbit horse (HRP) antibody and developed using the SuperSignal West Pico Chemiluminescent substrate. The arrowhead marks degradation products of CpxA. Click here to view larger image.
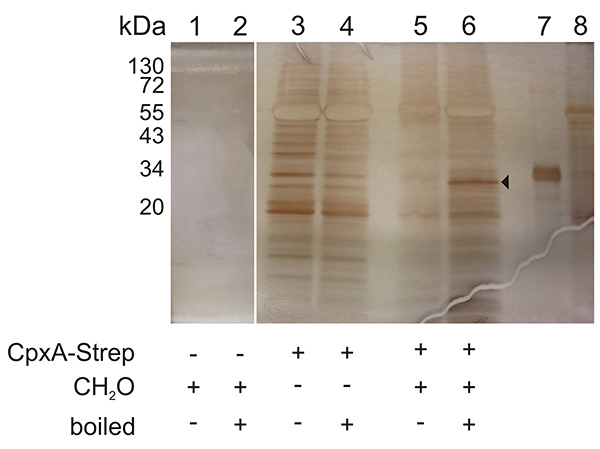
Figure 3. Representative silver-stained SDS-PAGE used for the identification of an interaction partner of a membrane protein by MS analysis. Samples corresponding to those of Figure 2 were silver stained. The arrow marks a band which was analyzed by MS analysis and which confirmed CpxR as the interaction partner of CpxA5. Lane 7 shows purified CpxR-His6 and lane 8 shows purified CpxA-His6 protein. Click here to view larger image.
