Three weeks after transplantation, the LN/fat pad chimera is recovered from the kidney. The chimera is now very similar to a normal LN in its own fat pad, and the LN is visible in the center of the adipose tissue (Figure 1). If the LN can't be identified, it is possible that it was lost during the transplantation on the kidney. When assessing the role of genes potentially important in LN development, the LNs recovered may remain very small and more difficult to find. This is the case when adipose tissue from LTβR-/- mice is reassociated with newborn LNs14.
Careful isolation of the LN allows further analysis of the progeny of EYFP+ adipose derived cells. Cryosections and immunofluorescence analysis of the LN reveal that EYFP+ adipose derived cells migrate into the LN where they contribute to the Gp38+ERTR-7+ LN stromal cell network (Figure 2A). Flow-cytometric analysis confirms that an important fraction of LN stromal cells derives from local EYFP+ adipose tissue progenitor cells, contributing to 30% of the CD45–Gp38+CD31– fibroblastic fraction and 10% of the CD45–Gp38–CD31+ blood endothelial cell fraction (Figure 2B). Up to 80% of the CD45–Gp38+CD31– fibroblastic fraction can derived from adipose tissue precursor cells demonstrating the crucial role played by adipose tissue in sustaining the growth of the LN stroma.
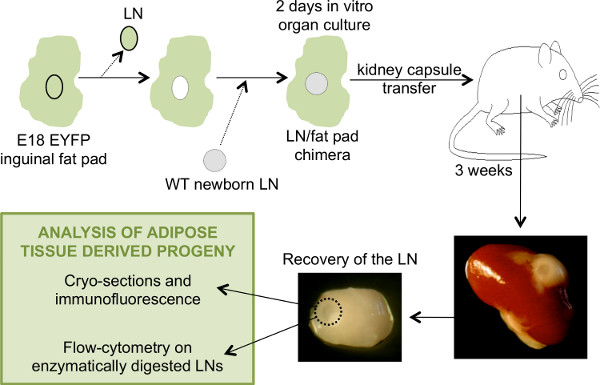
Figure 1. General schematic of the steps involved in the generation of LN/fat pads chimeras and the analysis of the LNs recovered after transplant. To generate a chimeric eYFP– LNs-eYFP+ fat pads, the LN from an E18.5 eYFP+ fat pad is removed and replaced by a WT eYFP– newborn LN. The chimeric LN-fat pad is placed in culture for two days before transplantation under the kidney capsule of an adult WT host mouse. After 3 weeks, the chimera is recovered and the LN isolated for analysis.
Click here to view larger image.
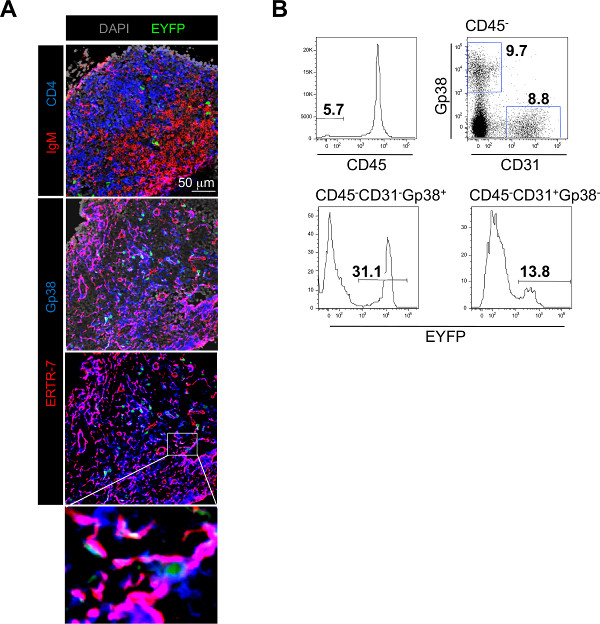
Figure 2. Typical experimental results obtained using the LN/fat pad chimera method. A) Immunofluorescence staining of frozen sections of a LN recovered from a LN-EYFP+ fat pad chimera showing the colonization of the LN by host derived IgM+ B cells in red and CD4+ T cells in blue and the presence of EYFP+ cells in green (top panel). The fibroblastic stromal cell network is stained with Gp38/Podoplanin in blue and ERTR-7 in red. EYFP+ cells derived from the fat pads in green are integrated to the stromal cell network (middle, bottom panels and enlargement). B) Flow-cytometric analysis of single cell suspension of enzymatically digested LNs recovered from LN-EYFP+ fat pad chimeras stained with CD45, CD31 and Gp38/Podoplanin. Percentage of CD45– stromal cells is shown in the histogram on the top row. Percentages of CD31–Gp38+ fibroblastic stromal cells and CD31+Gp38– blood endothelial cells in the CD45– fraction are shown in the dot plot on the top row. Percentages of fat pad derived-EYFP+ cells in the CD31–Gp38+ fibroblastic stromal cell fraction (bottom row, left histogram) and in the CD31+Gp38– blood endothelial cell fraction (bottom row, right histogram) are shown. Click here to view larger figure.
Click here to view larger image.
