Upon infection of tissue culture cells in vitro, S. flexneri can escape from the phagosome and invade the cytosol. In the cytosol, host cells can prevent the actin-based motility of Shigella by compartmentalizing bacteria inside septin cages (Figure 1A). Bacteria entrapped by septin cages can also be labeled by autophagy markers p62 (Figure 1B) and LC3 (Figure 1C). These observations highlight a novel mechanism of host defense that restricts dissemination of invasive pathogens, and also reveal new links between autophagy and the cytoskeleton. Strikingly, the depletion of autophagy markers significantly reduces septin caging of bacteria (Figure 2A), and work has also shown that the depletion of septin caging significantly reduces recruitment of autophagy markers8. Thus, at least in the case of Shigella, septin cage assembly and autophagosome formation can be viewed as interdependent processes. Other cellular requirements for compartmentalization of Shigella by septin cages include actin polymerization and actomyosin activity (Figure 2B).
There is no natural mouse model of shigellosis, and investigation of Shigella pathogenesis, septin biology and bacterial autophagy in vivo can benefit from a new animal model of infection, the zebrafish larvae19. It is possible to infect zebrafish larvae by injecting bacteria in various anatomical sites such as caudal intravenous injections for survival experiments, and tail muscle injections for in vivo microscopy (Figure 3A). Depending on the dose, S. flexneri injected in zebrafish larvae can either be cleared within 48 hr post-infection, or may result in a progressive and ultimately fatal infection (Figures 3B–3D). Shigella virulence factors are expressed at 28 ° C, the optimal growth temperature of zebrafish, and zebrafish infection by Shigella is strictly dependent upon its type III secretion system (T3SS)19, an essential virulence determinant in human disease. Taken together, these observations indicate that the zebrafish larva represents a valuable new host for in vivo analysis of Shigella infection.
The optical accessibility of zebrafish larvae enables visualization of septin caging in vivo (Figure 4A), an achievement that has never before been accomplished using mammalian host models. To complement evidence that septin cages entrap bacteria targeted to autophagy in vivo, one can infect transgenic zebrafish larvae expressing GFP-Lc3 and observe autophagy marker recruitment to Shigella (Figure 4B). For ultrastrucutral analysis of Shigella autophagosomes in vivo, electron microscopy can be used to clearly show the cytosolic sequestration of bacteria by double membrane vacuoles19. Autophagy is viewed as a key component of cell-autonomous immunity and a crucial defense mechanism against intracytosolic bacteria14-16. To characterize autophagy function in vivo, p62 morpholino-treated zebrafish larvae can be used. Unlike the core autophagy machinery [e.g., the 36 autophagy related proteins (ATGs)26], p62 is not essential for vertebrate development27 and thus zebrafish larvae can develop normally prior to infection. Strikingly, p62-depleted larvae inoculated with S. flexneri result in significantly increased mortality and increased bacterial burden19. In agreement with in vitro work showing that septin cage assembly is interdependent with autophagosome formation7,8, septin recruitment to Shigella is clearly reduced in p62-depleted larvae (Figure 4C). These data demonstrate that zebrafish survival depends on p62-mediated autophagy to control intracellular bacterial infection.
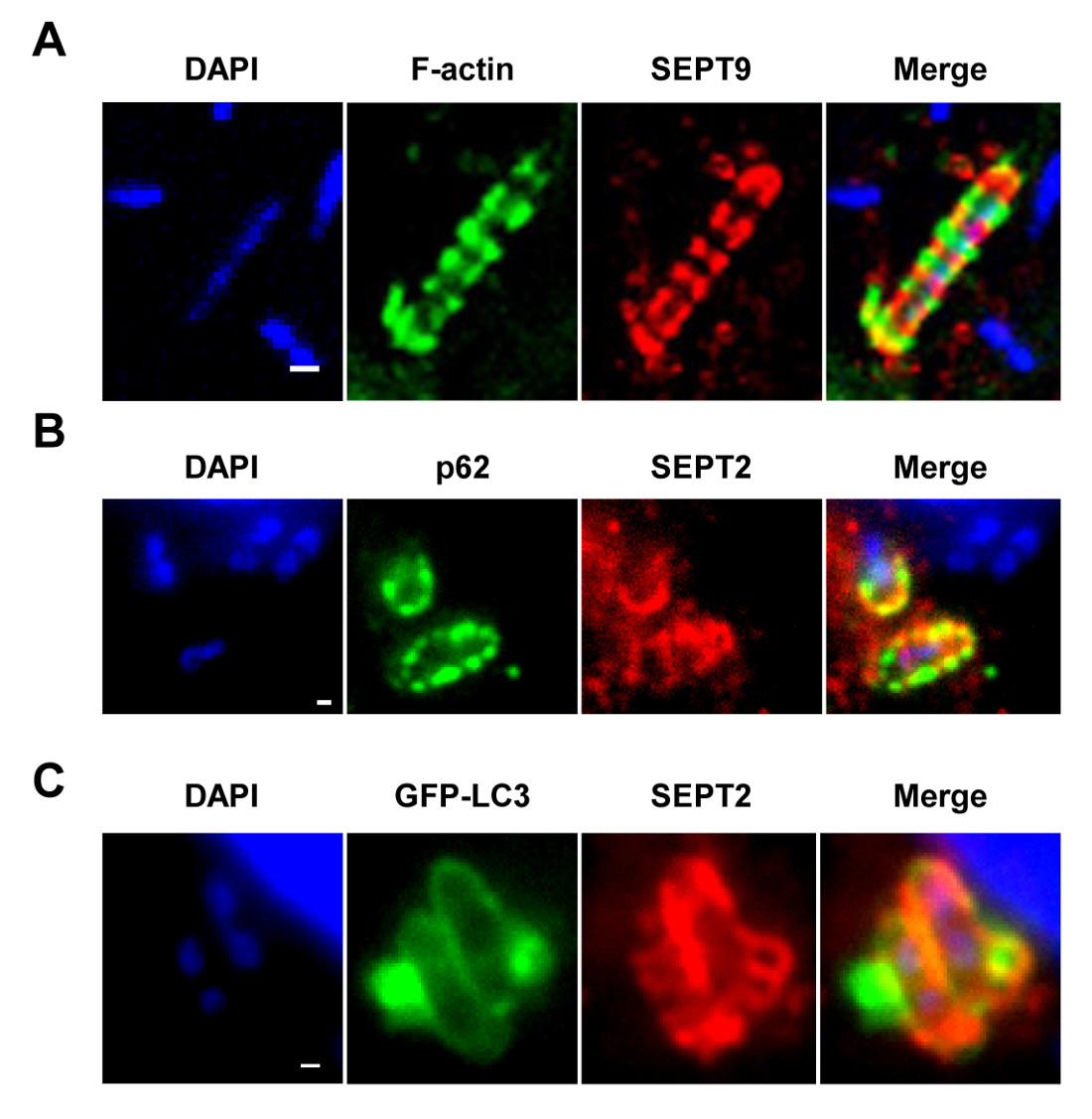
Figure 1. The septin cage in vitro. (A) HeLa cells were infected with S. flexneri for 4 hr 40 min, fixed, labeled with antibodies to SEPT9 and phalloidin, and imaged by confocal microcopy. Scale bar, 1 µm. (B) HeLa cells were infected with S. flexneri for 4 hr 40 min, fixed, labeled with antibodies to p62 and SEPT2, and imaged by fluorescent light microscopy. Scale bar, 1 µm. (C) HeLa cells were transfected with GFP-ATG8/LC3, infected with S. flexneri for 4 hr 40 min, fixed, labeled with antibodies to SEPT2, and imaged by fluorescent light microscopy. Scale bar, 1 µm. These figures have been modified from Mostowy et al7.
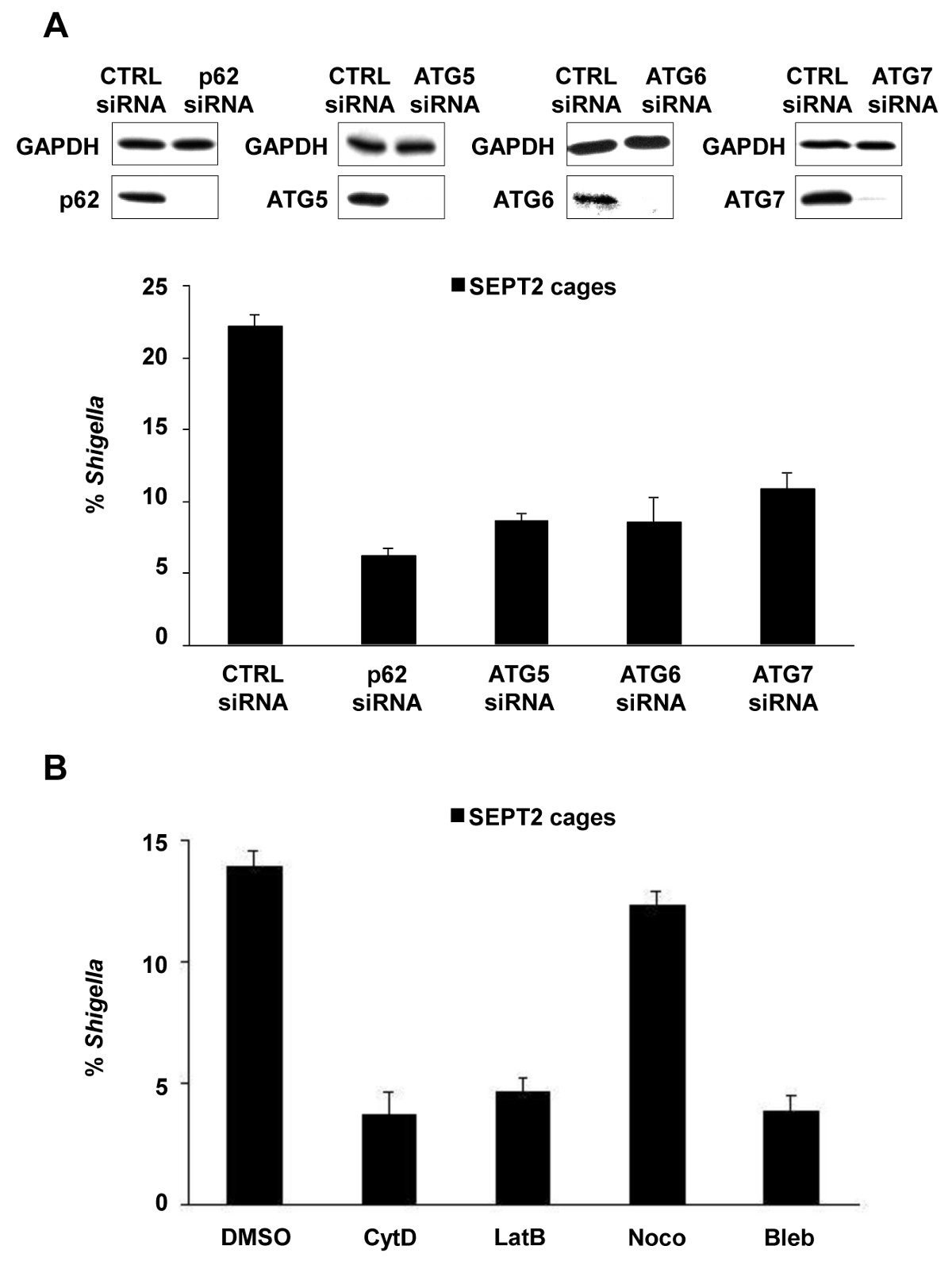
Figure 2. Cellular requirements for Shigella-septin cage formation. (A) HeLa cells were treated with control (CTRL), p62, ATG5, ATG6 or ATG7 siRNA. Whole-cell lysates of siRNA-treated cells were immunoblotted for GAPDH, p62, ATG5, ATG6, or ATG7 to show the efficiency of siRNA depletion (top). siRNA-treated cells were infected with S. flexneri for 4 hr 40 min, fixed, and labeled for quantitative microscopy. Graphs (bottom) represent the mean % ± SEM of Shigella inside SEPT2 cages from n ≥3 experiments per treatment. (B) HeLa cells were infected with S. flexneri, treated with DMSO, cytochalasin D (CytD), latrunculin B (LatB), nocodazole (Noco), or blebbistatin (Bleb) and after 4 hr 40 min were fixed and labeled for quantitative microscopy. Graphs represent the mean % ± SEM of Shigella inside SEPT2 cages from two independent experiments per treatment. These figures have been modified from Mostowy et al7.
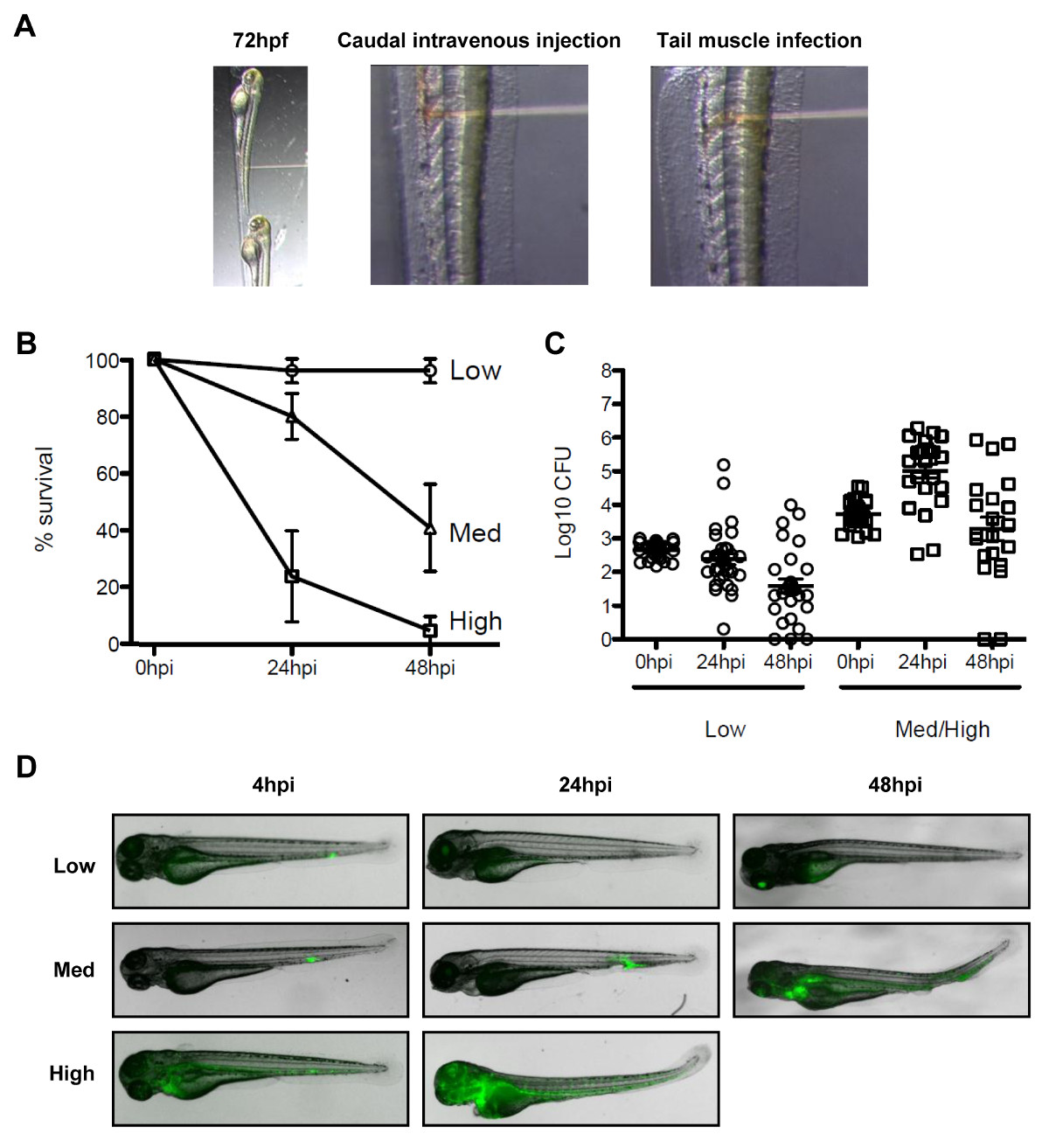
Figure 3. The zebrafish model of Shigella infection. (A) Images to illustrate orientation of the zebrafish larva under the stereomicroscope. (Left panel) Zebrafish larvae 72 hr post fertilization were positioned laterally in the injection plate with their dorsal side facing the injection needle. (Middle panel) Bloodstream infection was performed by injecting the bacteria (red solution) in the caudal vein, posterior to the urogenital opening. (Right panel) Infection in the tail muscle was performed by injecting the bacteria (red solution) over a somite. (B) Survival curves of 72 hr post fertilization larvae injected with various doses of S. flexneri and incubated at 28 °C for 48 hr post infection. The effective inoculum was classified as low (<103 CFU, open circles), medium (~4 x 103 CFU, open triangles) or high (~104 CFU, open squares). Mean % ± SEM (horizontal bars) from n ≥3 experiments per inoculum class. (C) Enumeration of live bacteria in homogenates from individual larvae at various times post infection measured by plating onto LB. Note, only larvae having survived the infection are included in enumeration analysis. Mean ± SEM (horizontal bars) also shown. (D) Distribution of GFP-Shigella determined by live imaging using a fluorescent stereomicroscope at various times post infection using a low, medium, or high dose inoculum (caudal intravenous injections). Overlay of transmission image (grey) and GFP fluorescence (green). (B)–(D)These figures have been modified from Mostowy et al19.
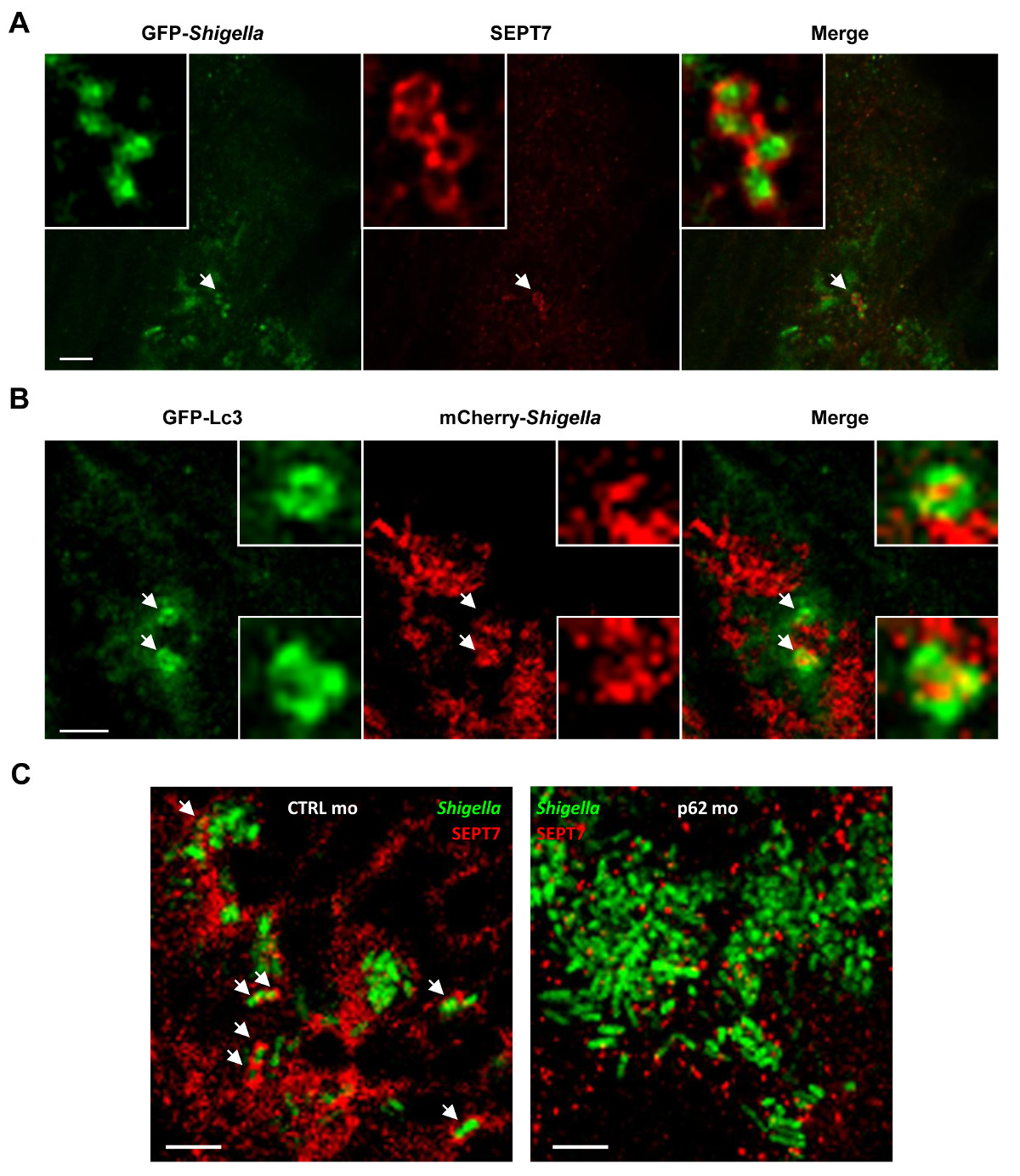
Figure 4. The cell biology of Shigella infection in vivo. (A) Zebrafish larvae were infected in the tail muscle with GFP-Shigella (low dose) for 24 hr, fixed, labeled with antibodies against SEPT7 (red) and to GFP (green), and imaged by confocal microscopy. Scale bar, 5 µm. (B) GFP-Lc3 zebrafish larvae were infected with mCherry-Shigella (medium dose) for 4 hr, fixed, labeled with antibodies against mCherry (red) and to GFP (green), and imaged by confocal microscopy. Scale bar, 1.5 µm. (C) Zebrafish larvae treated with either control (CTRL; left image) or p62 (right image) morpholinos were infected with GFP-Shigella for 4 hr (medium dose), fixed, labeled with antibodies against SEPT7 (red) and to GFP (green), and imaged by confocal microscopy. Arrows highlight some examples of Shigella entrapped in septin cages (CTRL) or not (p62 depleted) a 4 hr post infection. Scale bar, 5 µm. These figures have been modified from Mostowy et al19.
