TrCel5A and TrCel5A-mCherry were expressed successfully in tobacco plants using the transient expression system (Figures 1A and 1B). Examination of the expression pattern of the TrCel5A-mCherry fusion protein revealed healthy leaves (Figure 1C), which showed widespread protein expression under green light (Figure 1D).
Extraction of the total soluble proteins was performed on tobacco plants which had leaves that contained the transiently expressed TrCel5A protein and SDS-PAGE showed a large variety of proteins were present (Figure 2A, lane 1). The total soluble protein sample from TrCel5A expressing plants and a total soluble protein sample derived from NTEPs, were incubated for 10 min at 55 °C. TrCel5A remains stable and active at 55 °C while many other (native tobacco) proteins are not stable at this temperature. Thus, many non-essential proteins were precipitated by this treatment, leaving a partially purified sample of TrCel5A. After this thermal-precipitation step, a strong protein band was observed at ~40 kDa (Figure 2A, lane 1). This is likely to be the catalytic domain (CD) of the TrCel5A, which has been shown previously to run at this size when expressed in plants4. The negative and positive control samples (Figure 2A, lanes 3 and 4, respectively) showed low concentrations of remaining thermo-tolerant tobacco proteins from NTEPs (negative control, Figure 2A, lane 3) and multiple bands indicating a mixture of T. reesei proteins (positive control, Figure 2A, lane 4).
The size of the TrCel5A was confirmed by antibody staining directed at the His-6tag incorporated into the TrCel5A C-terminus, which was seen to be absent for both controls (Figure 2B). The TrCel5A was demonstrated to retain endoglucanase activity by CMC zymography (Figure 2C, lanes 1 and 2), where the refolded protein created a strong cleared area, indicating CMC degradation, at the same height as in the Coomassie stained SDS-PAGE gel and the Western blot. No activity was observed for the non-cellulase protein mixtures from NTEPs (Figure 2C, lane 3), and the mixture of T. reesei proteins run as a positive control showed endoglucanase activity from multiple components (Figure 2C, lane 4).
To quantify the cellulolytic activity of TrCel5A three assays were used. Firstly, the degradation of 4-MUC was analyzed by observing the increased fluorescence emission at 465 nm (excited at 360 nm) of the released 4-MU. TrCel5A was shown to degrade 4MUC, as did a 1:1,000 dilution of a commercially available mixture of T. reesei cellulases (Figure 3A).
A quantifiable analysis of the activity of TrCel5A against CMC was done by measuring the release of an Azo-dye at 590 nm from Azo-CMC. TrCel5A activity under these conditions, was equivalent in activity to a 1:1,000 dilution of the cellulase mixture mentioned above (Figure 3B).
The ability of TrCel5A to degrade filter paper was also analyzed. For this assay, an initial saccharification assay was performed (i.e. with the TrCel5A solution degrading the filter paper) and then the supernatant was analyzed by the PAHBAH assay to ascertain the quantity of reducing sugars released (Figure 3B).
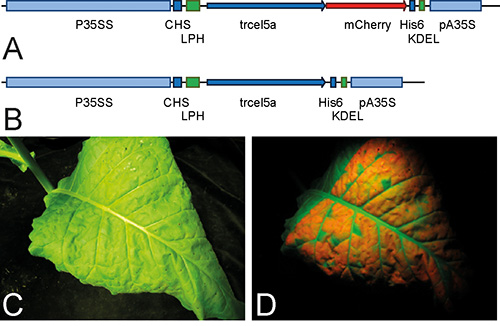
Figure 1. Plant expression cassettes for TrCel5A constructs and heterologous expression of TrCel5A-mCherry in tobacco leaves visualized by fluorescence under green light. Schematic representations of the plant expression cassette used for TrCel5A (A) and TrCel5A-mCherry (B) are shown. After transient TrCel5A-mC expression, tobacco leaves were examined under normal (C) or green light (D). For panels (A) and (B) the CaMV promoter (P35SS) and terminator signal (pA35S) are indicated in light blue. 5'-UTR of chalcone synthase (CHS), and the His6 coding sequence (His6) are indicated in blue. The plant codon-optimized leader peptide (LPH) derived from the heavy chain of the murine mAb24 is depicted in green. LPH achieves the secretion of the recombinant protein to the apoplast, after which a C-terminal KDEL signal, indicated in red, retards the protein to the ER. Arrows label the binding site for primer to amplify the CaMV 35SS expression cassette. For panels (C) and (D) the green light had an excitation 515 nm. Images were taken without magnification.
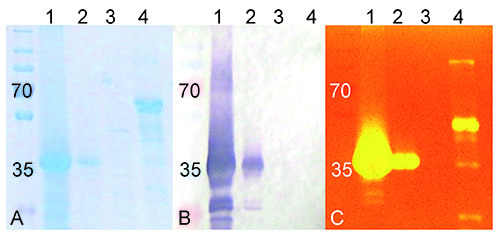
Figure 2. Size and activity of TrCel5A shown by SDS-PAGE, Western blotting, and CMC zymography. Separation of the total soluble proteins: from tobacco plants where TrCel5A was transiently expressed before (1) and after (2) thermal precipitation purification; from tobacco plants where TrCel5A was not present, also after thermal precipitation (3); or of a commercially available mixture of T. reesei cellulases (4). SDS-PAGE separated proteins are visualized with Coomassie Blue staining (A), Western blotting against the His-tag region of TrCel5A (B), and the cellulase activity shown by zymography using CMC visualized using Congo Red (C). PageRuler Plus (Fermentas) was used as the molecular weight marker (M).
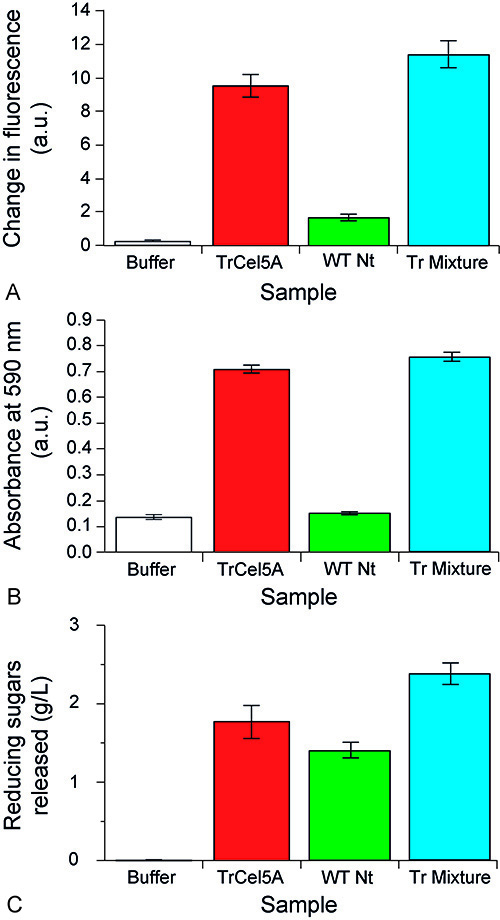
Figure 3. Enzyme activity quantification of TrCel5A. TrCel5A was incubated with 4-MUC (A), Azo-CMC (B), and filter paper (C) and the substrate degradation measured by release of the fluorophore 4-MU (A), an Azo-dye (B), or by quantifying reducing sugars by monitoring the color change of PAHBAH (C). Each assay shows the results for three replicates. Error bars indicate standard deviation. In all three assays the samples analyzed were buffer alone, TrCel5A after thermal precipitation purification, NTEP (WT Nt) after thermal precipitation purification, and a commercially available mixture of T. reesei cellulases.
