Using the general procedure described above, we adapted the protocol to assay the impact of soluble CD4 (sCD4) and coexpressed cellular CD4 on the exposure of CD4i epitopes on either wild-type (wt) or mutated Env, as described previously18,24,25,28. Figure 1 schematically represents the general procedure and the exposure of CD4i epitopes following treatment with sCD4 or by coexpression of cellular CD418. In Figure 2, we used sCD4 to induce Env conformational changes that expose CD4i mAbs 17b and 48d epitopes which overlap the coreceptor binding site24,29, whereas the outer-domain recognizing mAb 2G12 is not affected by this treatment as expected18,24.
The impact of point mutations in Env conformation can also be assessed using this assay, as presented in Figure 3. Here we used either the layer 1 Env mutant H66A, known to have a decreased propensity to spontaneously sample the CD4-bound conformation18,24,30,31 or a mutant (S375W) which predisposes Env to the CD4-bound state32 and obtained concordant results (Figure 3A). In cases where different Env expressors are used, it is often necessary to normalize the raw data expressed as relative light units (RLU) according to expression levels. In this case, we used PGT121, a mAb recognizing part of the Env glycan shield33-35, as the normalizing antibody (Figure 3B).
As we recently described18, interaction of Env and CD4 in the same cell leads to Env conformational changes that expose CD4i epitopes. In Figure 4, we cotransfected increasing amounts of a CD4 expressor together with Env in the cell-based ELISA assay and obtained increasing signals for CD4i mAbs A32 and C1118,36-38, which recognize discontinuous epitopes in the inner domain of gp120, whereas Env recognition by the conformational-independent 2G12 antibody was not affected (Figure 4A). In order to control for transfection efficiency between conditions, raw data was normalized to 2G12 (Figure 4B). Increased signals obtained for A32 and C11 antibodies depended on Env-CD4 interaction as indicated by the absence of A32 and C11 modulation when Env was cotransfected with a CD4 mutant (F43H) with decreased ability to interact with Env39.
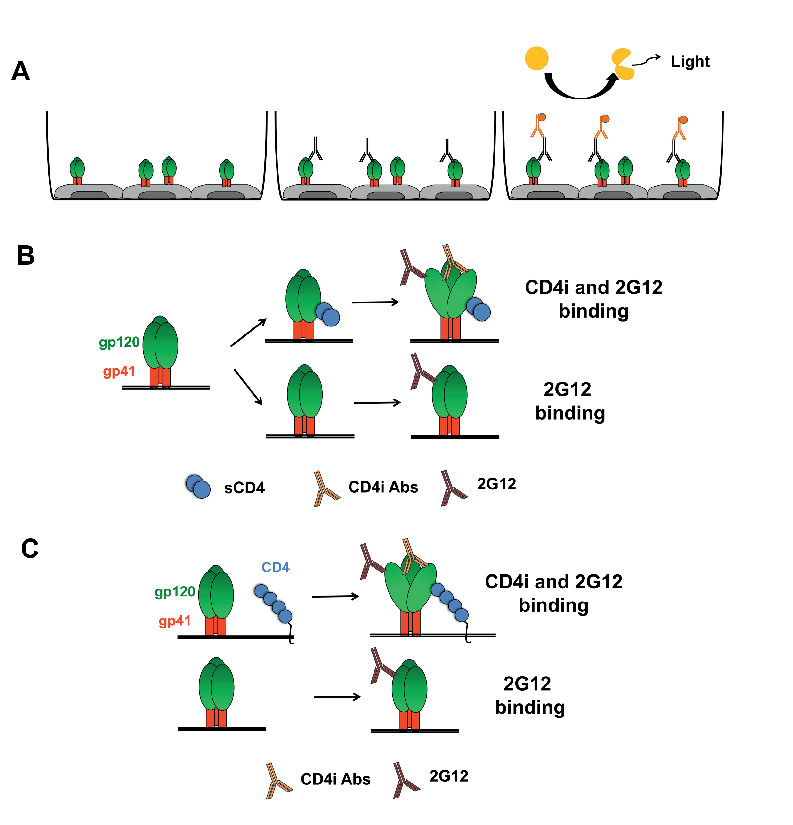
Figure 1. Schematic representation of the anti-Env cell-based ELISA. (A) General scheme of the procedure in which HOS cells are transfected to express trimeric Env at the cell surface. Env conformation can then be sampled by using different antibodies recognizing specific conformations (such as CD4i mAbs). Signals are detected by chemiluminescence after staining with HRP-conjugated anti-human mAbs. sCD4 (B) or coexpression of cellular CD4 (C) can be used to induce Env conformational changes that lead to exposure of CD4i epitopes.
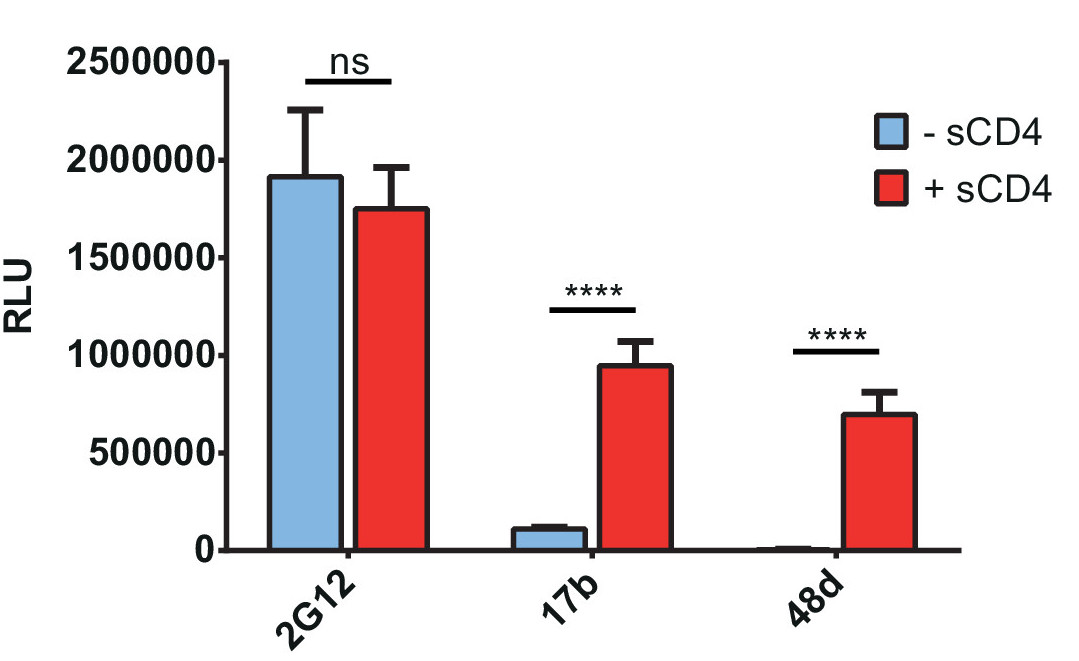
Figure 2. sCD4 induces Env conformational changes leading to exposure of CD4i epitopes. Interaction of sCD4 with HIV-1CO-JRFLΔCT Env enhances recognition by antibodies targeting CD4i epitopes (17b, 48d), but not by the gp120 outer-domain recognizing antibody 2G12. A plasmid encoding HIV-1CO-JRFLΔCT Env was transfected in each well and 48 hr later the cells were washed and incubated in presence or absence of 4 µg/ml sCD4 for 30 min at RT before continuing with the standard protocol (Day 4 – ELISA). Env conformation was then probed by incubating with 0.25 µg/ml 2G12, 1 µg/ml 17b or 48d anti-Env mAbs for 1 hr at RT. Signals were detected by chemiluminescence after incubation with an HRP-conjugated anti-human antibody for 45 min at RT. Shown are the mean RLU values ± SD of six replicates with signal obtained from wells transfected with an irrelevant plasmid (no Env) subtracted. Data is representative of results obtained in three independent experiments, with significance tested by two-way ANOVA (ns, not significant; ****, p <0.0001).
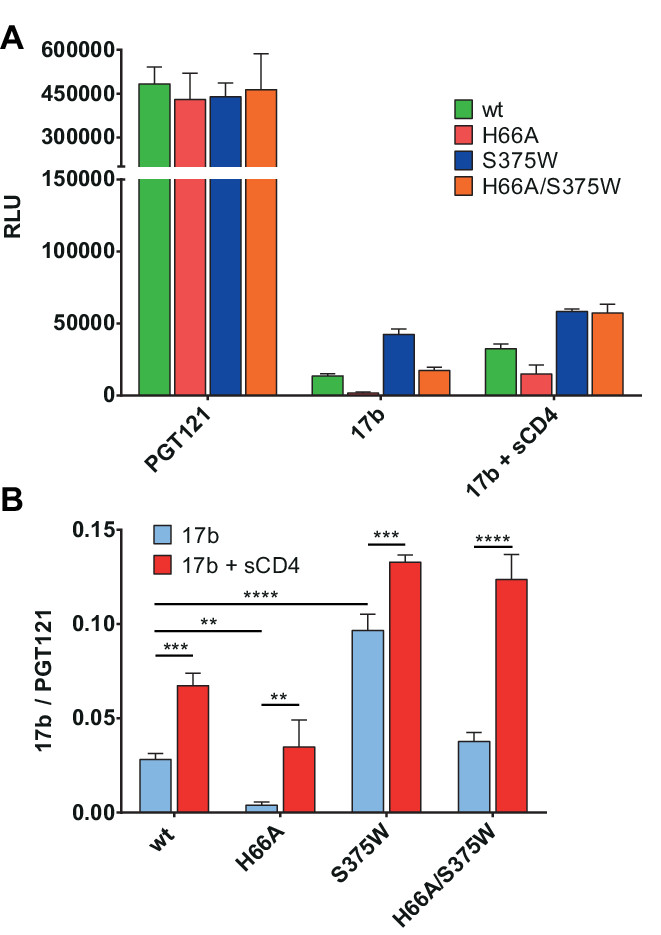
Figure 3. Modulation of Env conformation. HIV-1 YU2ΔCT layer 1 Env mutant (H66A) diminishes CD4i 17b recognition whereas the S375W variant exhibits increased 17b signal and is sufficient to restore the phenotype of the layer 1 mutant. (A) RLU values of the signals obtained using anti-Env PGT121 and 17b mAbs. (B) PGT121-normalized signals of CD4i mAb 17b following treatment with or without sCD4. Shown are the mean values ± SD of triplicates with signal obtained from wells transfected with an irrelevant plasmid (no Env) subtracted. Data is representative of results obtained in three independent experiments, with significance tested by two-way ANOVA (**, p <0.01; ***, p <0.001; ****, p <0.0001).
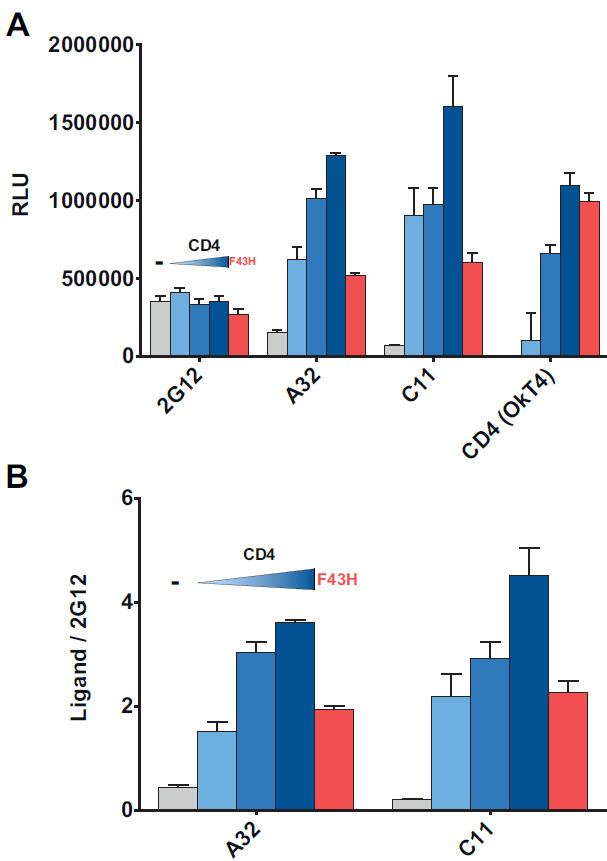
Figure 4. Coexpression of cellular CD4 enhances recognition by CD4i antibodies. A CD4-encoding plasmid was cotransfected with HIV1YU2ΔCT Env in order to favor the CD4-bound conformation18. (A) RLU values of the signals obtained using anti-Env 2G12, A32 or C11 mAbs and the anti-CD4 OkT4 mAb. (B) 2G12-normalized signals of CD4i mAbs A32 and C11. Shown are the mean values ± SD of triplicates with signal obtained from wells transfected with an irrelevant plasmid (no Env) subtracted. Data is representative of results obtained in three independent experiments. Grey bar indicates in absence of CD4, whereas the increasing blue bar indicates a step-wise increase in the amount of CD4 expressor being transfected (1.7 ng, 3.5 ng, and 7 ng) and red bar indicates the transfection of a CD4 mutant (F43H, 7 ng) with decreased capacity to interact with gp120.
