Mechanical vessel injury was performed on 2 dpf embryos (Figure 2A–C). Injury results in a rapid and reliable coagulation response as measured by time to cessation of bleeding (Figure 2D). To determine whether or not differences in the coagulation response could be measured, the anticoagulant hirudin was administered to the embryos by injection into the Duct of Cuvier immediately prior to wounding (5-10 nl of 1 unit per µl hirudin dissolved in water)(for demonstration of injections into the Duct of Cuvier, see previous JoVE article23)24. The administration of hirudin prior to injury resulted in significantly increased bleeding times versus vehicle control (Figure 2D).
Evidence of vessel damage and coagulation can be seen immediately post-injury using transgenic lines for endothelial (kdrl:egfp) and red blood cell (gata1a:dsred) markers25,26. Images were acquired sequentially every 5 min for a 12 hr period using epifluorescence. Representative still images are shown throughout different stages of wound repair (Figure 3). Using a combination of differential interference contrast (DIC) and fluorescence microscopy, it is possible to measure distinct parameters of wound repair. In order to determine whether or not wound repair followed a reproducible pattern across experiments, the time to reestablished blood flow was measured in 4 groups of fish. Vessel injury resulted in a reliable stereotypical response of 253 ± 16 min to the reestablishment of blood flow through the wounded vessel (n = 4-5 fish per experiment, average ± SEM).
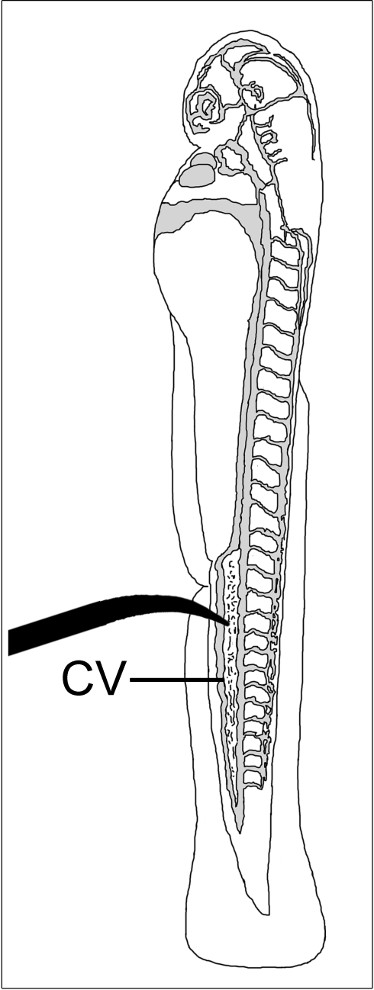
Figure 1: Diagram of 2 dpf embryo showing placement of minutia pin for performing mechanical injury of the caudal vein (CV). Vascular compartment is shaded in grey.
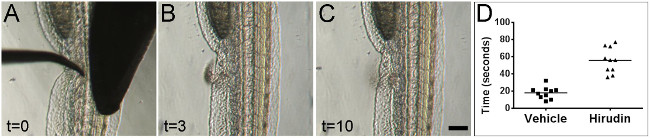
Figure 2: Bleeding times can be visually measured after mechanical injury. Stills from real-time video of zebrafish vessel injury on 2 dpf embryos. Images are shown at the time of injury (A), during active blood loss from the wound (B), and after cessation of blood loss (C). All times indicated are in seconds. Embryos are oriented laterally with anterior at top and ventral surface facing to the left. Scale bar 100 µm. Administration of the anticoagulant hirudin led to significantly increased bleeding times versus vehicle control (D) (p <0.0001, Student’s t-test).Please click here to view a larger version of this figure.
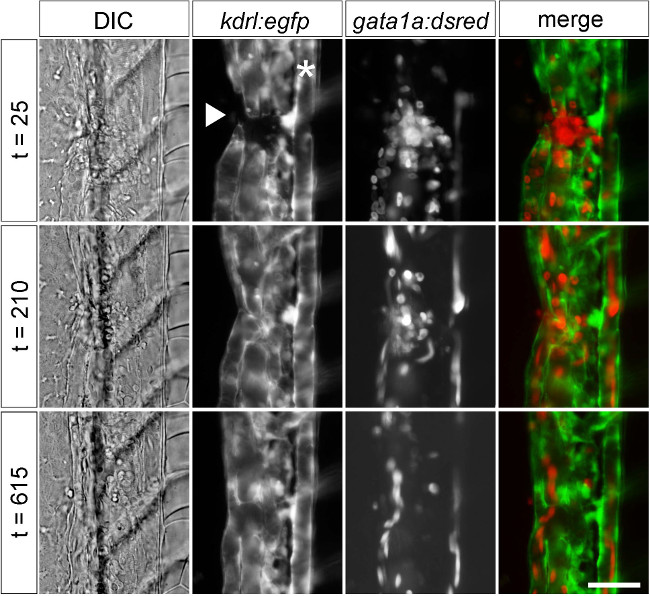
Figure 3: Visualizing mechanical injury and repair using transgenic markers. Stills from timelapse DIC and fluorescence microscopy after vessel injury using markers for vascular endothelium (kdrl:egfp) and red blood cells (gata1a:dsred) in 2 dpf embryos. Images showed a gap in vessels and local red blood cell accumulation (t = 25), partial repair with re-established blood flow (t = 210), and apparently complete restoration of normal vessel structure (t = 615). Time is indicated in minutes. Embryos are oriented laterally with anterior at top and ventral surface facing to the left. * indicates the position of the dorsal aorta. The injury (arrowhead) disrupted the caudal vein and part of the caudal plexus. Scale bar 25 µm.
