The data shown in the figures here were recorded in the unfolded hippocampus preparation with 4-AP (100 µM) aCSF added during incubation of the tissue in the recording chamber at RT (25 °C). Normally activity starts within 5 min, but in some hippocampal tissues from the older animals it may take longer. The 4-AP-induced neuronal firing observed with the PMEA is the same as previously reported 14,15. Since the electrodes have a height of 200 µm, the electrode tips are located just below the cell layer (Figure 3C) because the cell layer is usually 250 to 300 µm above the alveus of the hippocampus(Figure 2), recordings (57 out of 64 in the sample experiment) from different channels have a mostly positive deflection. However, some of these positive recordings could display small negative deflections as well (Figure 5B) if the recording electrode tips are close enough to the cell layer. If the electrode tips are located right at the level of cell layer, the recording will have very sharp negative spiking with positive shift on it or just negative spiking (Figure 5C) 16. In the data sample shown here, 5 channels out of 57 recordings have negative spiking. After acquiring the data from all the 64 channels, the method of individual normalization (Figure 6B) is applied to map the neural propagation on a 2-D plane along the time axis of the recording 7. With a combination of the PMEA and the unfolded hippocampus, neural propagation is mapped and observed to initiate mostly in one side of CA3 and move longitudinally in a diagonal wave front, crossing the entire area of the hippocampus (Figure 6).
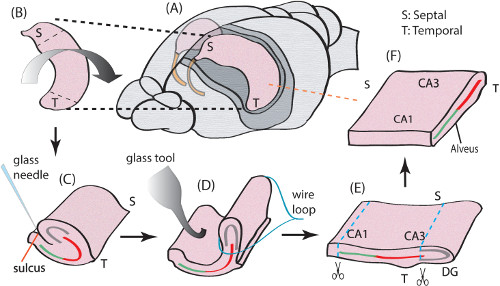
Figure 1. Surgical procedure for an unfolded hippocampus. (A) One of the two hippocampi is dissected from temporal lobe of a mouse brain. (B) Septal and temporal terminations along the longitudinal axis. (C) The hippocampus is flipped over with a fire polished glass pipette to expose the sulcus. Both ends of the hippocampus are trimmed and a glass needle is used to cut the connections between DG and CA1 or subiculum along the longitudinal axis. (D) The hippocampus is unfolded by a custom-made metal wire loop. (E) Unfolded hippocampus with subiculum and DG trimmed. (F) The final tissue preparation of an unfolded hippocampus. This position shows the orientation of the hippocampus when it is placed on the array in an experiment. For additional details about the unfolded hippocampus anatomy please refer to experimental methods in previously published manuscripts 7. Please click here to view a larger version of this figure.
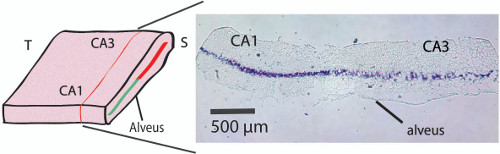
Figure 2. Histology cross-section of the unfolded hippocampus. Crystal-violet stain shows the position of the CA1-CA3 pyramidal cell layer within the unfolded hippocampus a level of 250 to 300 µm above the alveus thereby locating the microelectrodes right below the cell layer (refer to Figure 3C). To obtain the sections stained with the crystal violet, the tissue was post-fixed after the hippocampus was unfolded. The unfolded hippocampus was placed in PFA 4% O/N. Then, the tissue was transferred and kept in sucrose solution (30%) for 48 hr, and followed by snap-freezing in a biker containing isopentane (2-Methylbutane) to cool down to -35 °C in dry ice. Frozen section were then cut in a cryostat with 20 µm thick sections in the transverse plane (as shown in the Figure 2) to reveal the location of the pyramidal cell layer. Please click here to view a larger version of this figure.
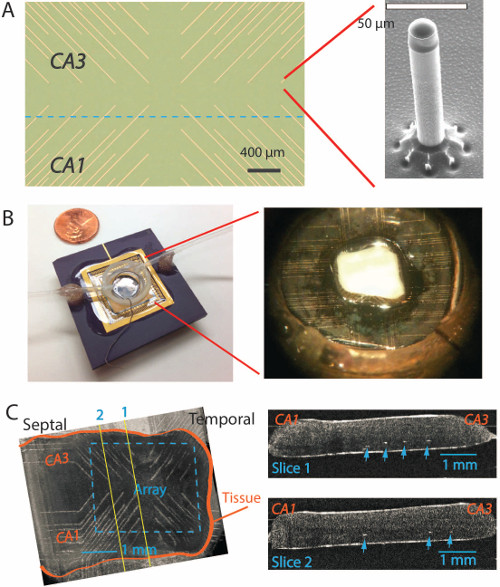
Figure 3. Unfolded hippocampus on the PMEA. (A) Top view of a new PMEA with the microelectrodes located at the end of each gold metal trace. The insert on the right side shows a single microelectrode under electron microscopy with a height of 200 µm and a diameter of 20 µm 7,13. (B) The microelectrode array is glued on a PGA package with a plastic recording chamber around the microelectrodes on a glass substrate. The insert on the right shows an unfolded hippocampus placed on the microelectrode array in the middle. (C) On the left, Optical Coherence Tomography(OCT) imaging shows the unfolded tissue positioned on top of the array in a different experiment. On the right, two longitudinal slice images obtained from the OCT imaging on the left shows the microelectrode tips (white dots pointed by the arrows) reach into the area right below the cell body layer. Please click here to view a larger version of this figure.

Figure 4. Experimental setup. (A) A plastic cover lid with screws is placed over the circuits to protect it against possible water damage. The recording chamber has both inlet and outlet tubes to carry the solution flow. A custom made tissue anchor glued with a Nylon fiber mesh is used to secure the tissue during the experiments. (B) Picture taken from the bottom of the glass substrate of the PMEA showing the tissue anchor holding a sample slice during an experiment. The curved wires are the Nylon fibers from the mesh pressing on top of the tissue. The round dots are the bases of microelectrodes. Arrows point to electrodes that were damaged after several experiments. Please click here to view a larger version of this figure.
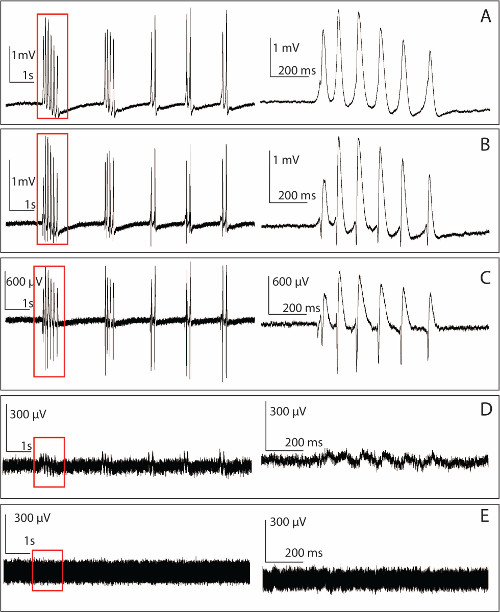
Figure 5. Raw data recorded by the PMEA from an unfolded hippocampus. Left panel: 4-AP-induced epileptiform activity recorded from a single microelectrode. Right panel: Zoomed-in version of the signal marked in the time window on the left. (A) 10 sec section of an example of the spontaneous activity induced by 100 µM 4-AP aCSF obtained for one of the microelectrodes located in the basal dendritic region based on the polarity of the signals with a SNR of 34.9 dB. (B) Raw data from another microelectrode located closer to the somata with a SNR of 27.2 dB. (C) Example of recording obtained from an electrode positioned within the somatic layer. In this example, the SNR is 18.53 dB. (D) Recording obtained from a bent electrode still conducting but not penetrating into the tissue. The bent electrode has a significant lower SNR compared to those intact electrodes (1.5 dB in this example). (E) Baseline noise recorded from a microelectrode. The baseline usually has a peak to peak value from 150 to 200 µV and the impedance of a single electrode is around 1 to 2 MΩ. Please click here to view a larger version of this figure.
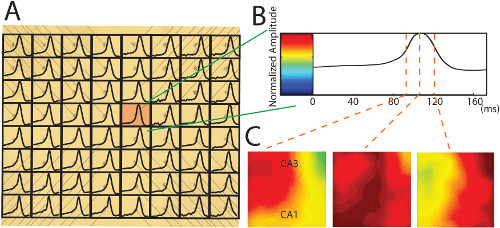
Figure 6. Conversion of neural recording data into the 2-D propagation maps. (A) A 170 msec time window truncates a single neural spiking in each channel plotted on the photo of the PMEA. The raw data is filtered by a low pass filter at 100 HZ. The signals from the broken electrodes are interpolated with the recordings around it. In this example, all the spikes are positive. (B) A single neural spike from the red pixel in (A) is normalized to the color bar with a custom-developed method for individual normalization (Please refer to the previous publication for more details about the normalization 7). (C) The color maps created by the individual normalization show that propagation moves across the entire area of the unfolded hippocampus at different time points. Please click here to view a larger version of this figure.
