The behavioral responses elicited from mice starting at 30 °C, 23 °C, 17 °C, and 12 °C are highly reproducible (Figure 4A)20. In order to measure the cold stimulus being generated under the hindpaw, mice were anesthetized with a ketamine/xylazine/acepromazine cocktail and their paws were secured on the glass on top of a T-type filament thermocouple (Figure 4B)20. The glass was cooled or warmed to the desired testing range. Although the plate is cooled uniformly along the length of the plate (Figure 5A)2, it should be noted that a cold gradient is generated across the behavior enclosures (Figure 5B)2. The parts of the enclosure that are closer to the dry ice on either side of the enclosures are cooler, while the central parts are slightly warmer (Figure 5B)2. In the coldest temperatures used, the mice spend the majority of their time in the central parts of the enclosure. Once the glass plate temperature had stabilized, a focal dry ice stimulus was placed on the glass underneath the paw/thermode. Based on the temperature traces recorded from this setup, it is clear that the cold stimuli generated using the CPA are highly reproducible at each temperature range (Figure 4C)20.
The cold stimulus generated in the CPA was also measured using three different thicknesses of glass to vary the intensity of the cooling (Figure 3). The rate of cooling is inversely related to the thickness of the glass, and any of these thicknesses can be used to measure cold sensitivity as needed (Figure 3).
Previous work has shown that the CPA can detect analgesia and hypersensitivity in mice. 30 min after subcutaneous injections of 1.5 mg/kg of morphine, mice have significantly longer latency to withdrawal than mice given subcutaneous injections of saline (Figure 6A: 2-way ANOVA main effect *p < 0.05 with Bonferroni post-hoc test; 30 min **p < 0.01; n = 12 per group)1. By 60 min after morphine/saline, there is no difference between the saline- and morphine-injected groups, which is consistent with the rate of morphine metabolism in mice.
Complete Freund’s Adjuvant (CFA) has previously been shown to cause inflammation and hypersensitivity after hindpaw injection21. After CFA injections, the CPA withdrawal latencies decrease 2 and 3 hr post-injection (Figure 6B: 2-way ANOVA main effect p < 0.001 with Bonferroni post-hoc test; 2 hr *p < 0.05, 3 hr **p < 0.01 n = 12 per group). 4 hr after CFA injection, the mice were given subcutaneous injections of 1.5 mg/kg morphine. 30 min after morphine injection, both CFA- and saline-injected mice had increased withdrawal latencies relative to their latencies at 3 hr (Figure 6B: 1-way ANOVA with Dunnett’s post-hoc test; CFA 3 hr vs. CFA 4.5 hr $$$p < 0.001, saline 3 hr vs. saline 4.5 hr $$$p < 0.001). An hour later, once the morphine had been metabolized, the CFA-injected mice once again had lower withdrawal latencies than the saline-injected control mice (Figure 6B: 2-way ANOVA with Bonferroni post-hoc test; **p < 0.01)1.
Most mammalian species have the ability to adapt their temperature sensitivity to match their environment. In vitro, studies have suggested that this adaptation process is dependent on PIP2 hydrolysis22-24, but previous behavioral tools were unable to validate this hypothesis in vivo. The CPA is capable of quantifying this adaptation in two different ways. By testing the withdrawal latency of mice as the glass cools (Figure 7A,C), the CPA can measure cold adaptation as it happens2. Under normal conditions the withdrawal latency is unchanged as the plate cools, suggesting that cold adaptation happens faster than can be quantified with the CPA (Figure 7B: 0 min = 12.13 ± 0.8 sec, 30 min = 12.1 ± 1.6 sec, 60 min = 13.2 ± 1.1 sec, 90 min = 10.8 ± 1.2 sec 1-way ANOVA with Bonferroni post hoc test p > 0.05, n = 6)2. However, when mice are given intraplantar injections of the phospholipase-C inhibitor U7312225 before the plate is cooled (Figure 7C) their withdrawal latencies are decreased, suggesting that adaptation is impaired (Figure 7D: baseline = 11.29 ± 0.53 sec, 30 min = 8.09 ± 1.17 sec; 1-way ANOVA with Dunnett’s post-hoc test, main effect p = 0.02, individual baseline vs. 30 min p = 0.02, n = 9).
The CPA can also measure the ability to adapt to cold ambient temperatures over long periods of time. When wild-type mice are tested using the CPA after being acclimated for 3 hr at 30 °C, 23 °C, 17 °C, or 12 °C the withdrawal latency is the same at all starting temperatures, suggesting that the wild-type mice adapted to the colder ambient temperature (Figure 2A: WT 30 °C = 13.23 ± 0.5 sec, 23 °C = 12.8 ± 0.7 sec, 17 °C = 12.3 ± 0.9 sec, 12 °C = 12.8 ± 0.5 sec, 1-way ANOVA with Bonferroni post-hoc test, p > 0.05 n = 6 for 30 °C, n = 15 for 23 °C, 17 °C, and 12 °C)20. Unlike the wild-type mice, as the starting temperature decreases the withdrawal latencies of TRPM8-KO mice decrease, suggesting that they are unable to adapt their response threshold to suit their environment (Figure 8: 1-way repeated measures ANOVA with Bonferroni post-hoc test; males main effect p = 1.5 x 10-5, 12 °C vs. 23 °C p = 6 x 10-5, 17 °C vs. 23 °C p = 0.004; females main effect p = 3.6 x 10-5, 12 °C vs. 23 °C p = 9.25 x 10-5, 17 °C vs. 23 °C p = 0.0005; df = 1, n = 11 males and 11 females)20.
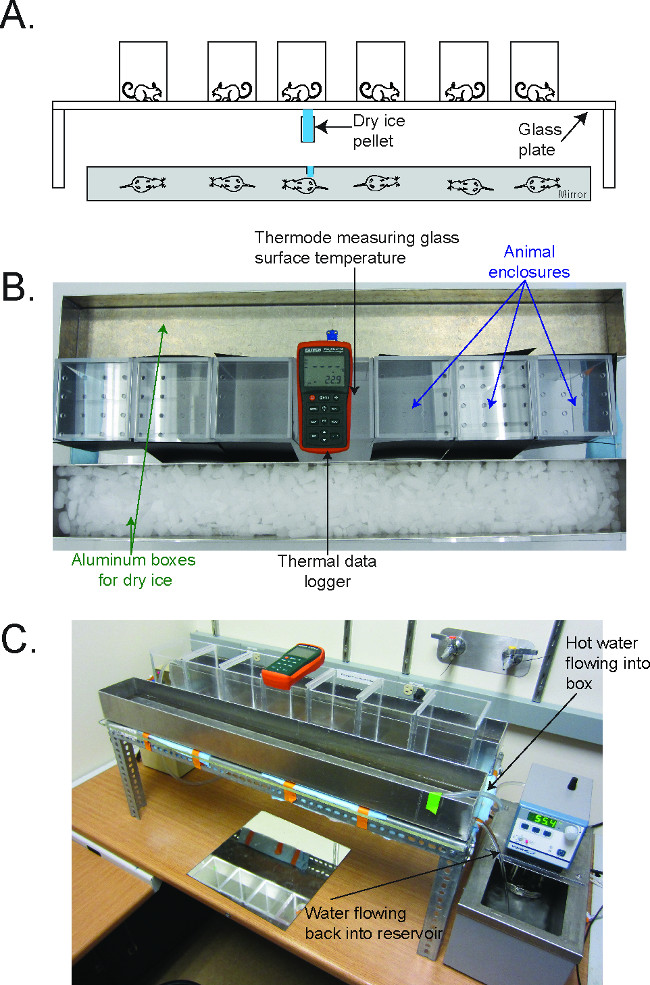
Figure 1. The Cold Plantar Assay (CPA) apparatus2. (A) Schematic for performing the CPA. Mice are acclimated on a glass plate in plastic behavioral enclosures until they are at rest. A dry ice pellet is applied to the underside of the glass underneath the hindpaw, and the latency to withdraw from the cooling glass is measured. (B) Picture of the CPA apparatus, in configuration to cool the plate to 5 °C. The thermal data logger is in the center of the enclosures, and the aluminum boxes flank the enclosure on either side. (C) Picture of the CPA apparatus, in configuration to warm the plate to 30 °C. The water circulator flows hot water into the aluminum box, which then flows out the drain on the side back into the reservoir of the circulator. Reused with permission from Brenner et al. 2014 2. Please click here to view a larger version of this figure.
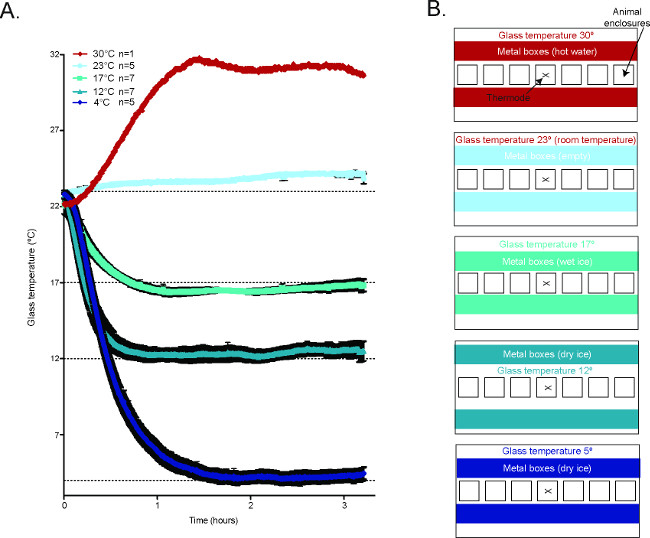
Figure 2. Temperature of the glass plate during the CPA2. (A) Averaged temperature tracings of the glass plate during behavioral experiments in the CPA. 30 °C n = 1, 23 °C n = 5, 17 °C n = 7, 12 °C n = 7, 4 °C n = 5. (B) Schematic diagrams demonstrating how to generate the different temperature conditions in the CPA. Reused with permission from Brenner et al. 2014 2. Please click here to view a larger version of this figure.
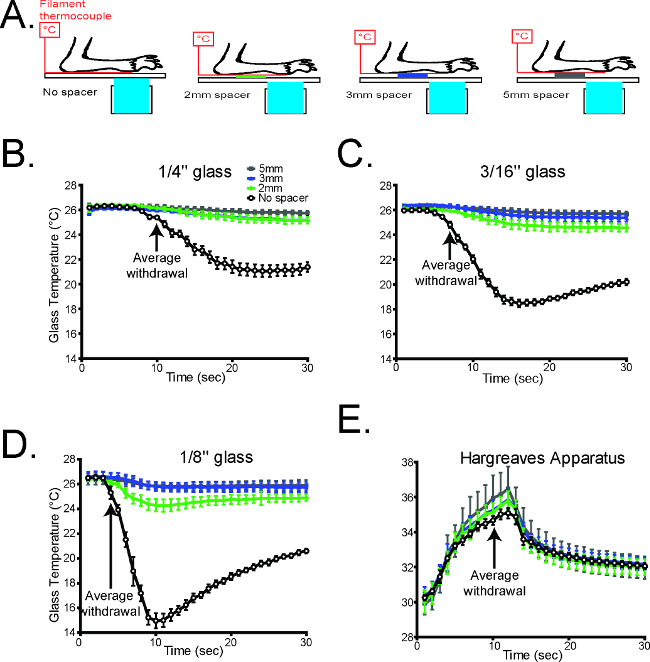
Figure 3. The glass thickness is inversely correlated with the rate of cooling1. (A) Schematic diagramming the experimental design (B-D). The temperature during cold plantar stimulus underneath the paw was measured on all three glass thicknesses under normal conditions, and which Styrofoam spacers propping the paw away from the glass surface. In all cases, propping the paw away from the glass caused a dramatic decrease in the cold stimulus measured at the paw (n = 6 per glass thickness). (E) The temperature underneath the paw was measured during Hargreaves radiant heat stimulation with the paw propped up with Styrofoam. Unlike the cold plantar assay, the thermal stimulus in the Hargreaves assay is largely unaltered when the paw is propped away from the glass (n=6). Reused with permission from Brenner et al. 20121 Please click here to view a larger version of this figure.
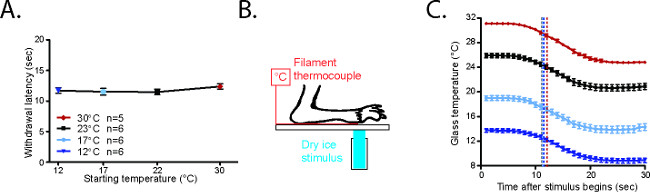
Figure 4. CPA withdrawal latencies are consistent20. (A) Average withdrawal latency for mice starting from 23 °C, 17 °C, or 12 °C. (B) Configuration to measure CPA cold stimulus. The paw of an anesthetized mouse is secured on the glass plate with laboratory tape on top of a T-type filament thermocouple. The CPA stimulus is placed on the underside of the glass underneath both paw and thermocouple. (C) Temperatures generated in the CPA starting from 30 °C, 23 °C, 17 °C, or 12 °C. The black arrows represent the average withdrawal latencies for awake mice in each condition. Reused with permission from Brenner et al. 20142. Please click here to view a larger version of this figure.
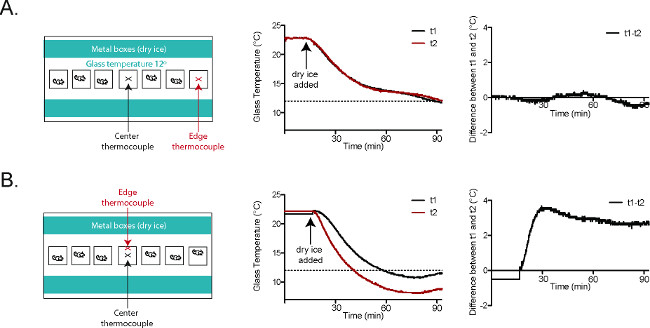
Figure 5. Glass plate temperatures are consistent in the CPA2. (A) Thermocouple t1 (black) was placed at the center of the plate. Thermocouple t2 (red) was placed in the behavioral enclosure closest to the right edge of the plate. The temperature tracings and the graph at the far right (t1-t2) show nearly identical temperatures at t1 and t2 throughout the course of the experiment. (B) Thermocouple t1 (black) was placed at the center of the plate. Thermocouple t2 (red) was placed in the central behavioral enclosure, at the wall closer to the dry ice-filled aluminum boxes. The temperature tracings and the graph at the far right (t1-t2) show that there is a roughly 3 °C difference between t1 and t2 once the plate has reached a stable temperature. Reused with permission from Brenner et al. 20142. Please click here to view a larger version of this figure.
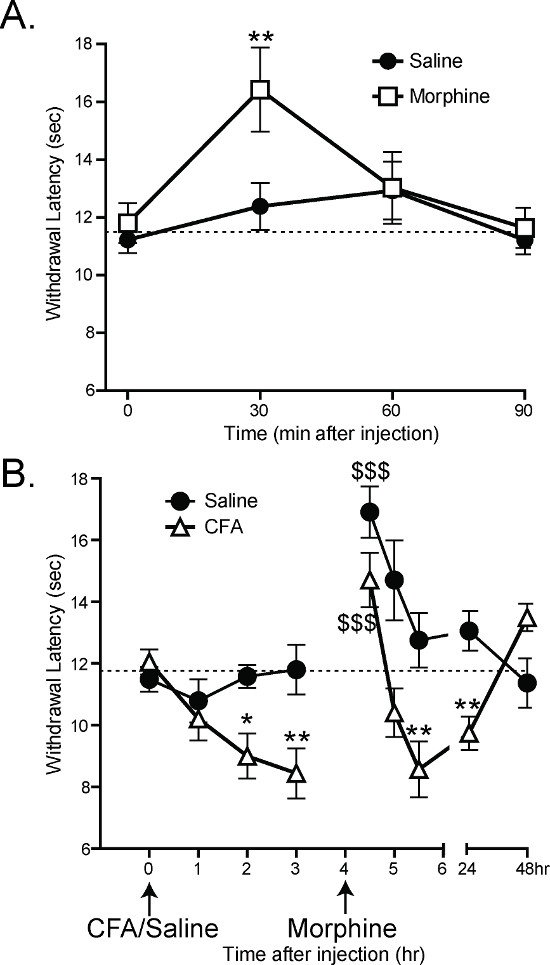
Figure 6. The CPA can measure analgesia and hypersensitivity1. (A) Subcutaneous injection of 1.5 mg/kg morphine increases the withdrawal latency of mice 30 min after injection (2-way ANOVA with Bonferroni post-hoc test; 30 min post injection **p < 0.01). 60 min after injection, there is no significant difference between morphine-injected and saline-injected mice. (B) Intraplantar injection of 10 ul Complete Freund’s Adjuvant (CFA) decreases the withdrawal latency of mice 2 and 3 hr post injection (2-way ANOVA with Bonferroni post-hoc test; *p<0.05, **p<0.01). All mice were given subcutaneous injections of morphine at 4 hr, and all withdrawal latencies at 4.5 hr were significantly higher compared with 3 hr (1-way ANOVA with Dunnet’s post-hoc test; $$$p<0.001). 5.5 hr after injection of CFA (1.5 hr after morphine injection), CFA-injected mice still had lower withdrawal latencies than saline-injected mice (2-way ANOVA with Bonferroni post-hoc test; **p<0.01). Reused with permission from Brenner et al. 20121. Please click here to view a larger version of this figure.
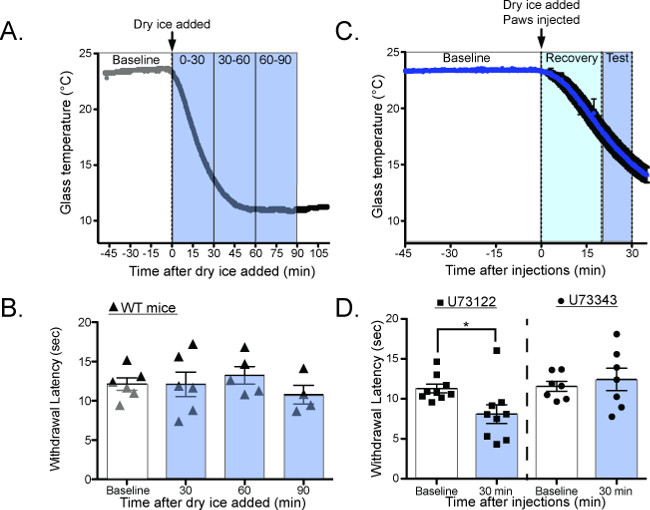
Figure 7. Measuring cold adaptation as the glass plate dynamically cools20. (A) Schematic for performing the CPA as the glass plate is cooling. Baseline temperatures are measured at RT, the dry ice containers are added to the plate, and the withdrawal latency is measured as the glass plate cools. (B) Wild-type mice have the same withdrawal latency as the glass plate cools, suggesting that they adapt to cooling temperatures faster than can be measured with the CPA (Baseline = 12.8 ± 0.3 sec, 30 min = 13.67 ± 0.9 sec, 60 min = 11.03 ± 1.0 sec, 90 min = 11.31 ± 0.6 sec, n = 3 mice; 1-way ANOVA with Bonferroni post-hoc test, no significant differences between any groups). (C) Schematic for performing the CPA as the glass plate is cooling, after intraplantar injections of the PLC inhibitor U73122 or the control compound U73343. (D) Mice have significantly lower withdrawal latencies while the plate is cooling after U73122 injection, suggesting that U73122 interferes with the ability to adapt to cooling ambient temperatures. Reused with permission from Brenner et al. 2014 20. This Figure has been reproduced with the permission of the International Association for the Study of Pain (IASP). The figure may NOT be reproduced for any other purpose without permission. Please click here to view a larger version of this figure.
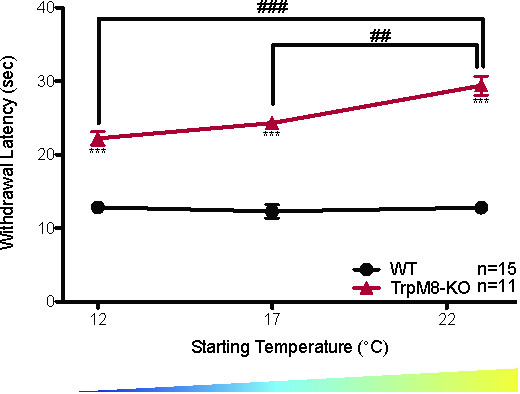
Figure 8. TRPM8-KO mice do not adapt to environmental cooling20. TRPM8-KO mice have higher withdrawal latencies than wild-type littermates at all starting temperatures measured (2-way ANOVA with Bonferroni post-hoc test; ***p < 0.001). The withdrawal latency of TRPM8-KO mice also decreases as the starting temperature decreases (1-way ANOVA with Bonferroni post-hoc test; ##p < 0.01, ###p < 0.001), while there is no significant change in the withdrawal latency of wild-type littermates as the starting temperature decreases. Reused with permission from Brenner et al. 2014 20. This Figure has been reproduced with the permission of the International Association for the Study of Pain (IASP). The figure may NOT be reproduced for any other purpose without permission. Please click here to view a larger version of this figure.
