bFGF Withdrawal Decreases Proliferation in IMOP Cells
To decrease the proliferative capacity of iMOP cells and initiate differentiation of iMOP cells, bFGF was withdrawn from the cultures. To confirm that growth factor withdrawal decreases proliferation, EdU incorporation was employed as a proliferation assay. The percentage of cells that incorporated EdU from otospheres cultured with iMOP culture media (containing bFGF) and otospheres cultured in sensory epithelia media (without bFGF) was compared. Otosphere cultures were pulsed with the nucleotide analog EdU, harvested and fixed. Incorporated EdU nucleotides were fluorescently labeled with Alexafluor 488 azide using click-chemistry. Cells from otospheres grown in the absence of bFGF showed decreased incorporation of EdU relative to cells cultured with bFGF (Figure 1A-F). As the size of the otosphere increased, it was increasingly difficult to visualize all the cells from the otosphere with an epifluorescent microscope. To address this, cells were dissociated and mounted so that they can be unambiguously visualized en mass. Even after dissociation and fixation, the cells re-aggregated. To maintain the cells in a dissociated state, detergents that prevented the cells from re-aggregating were tested. Tween 20 was one of the detergents that prevented re-aggregation of the cells. Dissociated cells were washed with 0.1% Tween 20 and mounted onto slides. EdU labeled cells were then visualized by epifluorescence microscopy (Figure 1G,H). Representative results from individual experiments (n = 5) showed a significant decrease in the percentage of EdU labeled cells from 48% to 19% after 3 days after bFGF withdrawal (p<0.005). These results confirm a dramatic drop in the percentage of S phase cells and proliferative potential of iMOP cells after bFGF withdrawal.
IMOP-derived Sensory Epithelia Express Cdkn1b and Show Morphological Changes
A timeline of iMOP sensory epithelia differentiation protocol is shown (Figure 2A). Otospheres from proliferative cultures were dissociated into single cells and allowed to recover for 3 days in iMOP culture media. This method helps enrich for proliferating cells and selects against post-mitotic cells in these starting cultures. After 3 days, newly formed otospheres were seeded in sensory epithelia differentiation media and allowed to undergo unguided differentiation for 10 days. Bright field images of typical cultures containing otospheres at different time points after seeding showed an initial decrease in size before otospheres start increasing in size (Figure 2B-E).
Since brightfield microscopy cannot reveal many of the changes that occur during differentiation of iMOP cells, immunostaining with molecular markers was used to highlight morphological and molecular features of the differentiated cells27. To determine if expression of Cdkn1b (p27KIP) can be used as a marker for differentiation, otospheres from proliferative or sensory epithelia differentiated iMOP cultures were compared. Fluorescent markers were visualized and captured by epifluorescence microscopy. Representative images of otospheres from proliferative cultures showed low expression of Cdkn1b outside the nuclei with almost no phalloidin staining (Figure 3 A-D). Otospheres cultured in sensory epithelia differentiation media displayed increased expression of nuclear Cdkn1b concomitant with the appearance of phalloidin labeling in the peripheral edges of cells (Figure 3 E-H). In proliferating iMOP otospheres, Cdh1 (E-cadherin) and phalloidin are weakly labeled (Figure 3 I-L). In differentiated iMOP otospheres, pronounced phalloidin and Cdh1 labeling highlight the morphological changes in actin filaments and regions of cell-cell adhesion (Figure 3 M-P), similar to previous results27. During sensory epithelia differentiation, the morphological changes in actin filaments parallel the appearance of Cdh1 expression in adhesion sites between cells. In this protocol, the increase in Cdkn1b expression along with the changes in phalloidin and Cdh1 serve as early indicators of sensory epithelia differentiation in iMOP cells.
IMOP-derived Neurons Express Cdkn1b and Tubb3
A schematic of iMOP neuronal differentiation protocol is shown (Figure 4A). Similar to the aforementioned protocol, iMOP cells were dissociated and allowed to recovery for 3 days in iMOP culture media. Otospheres were harvested, dissociated into single cells and seeded onto poly-D-lysine and laminin coated coverslips. The cells were allowed to differentiate for 7 days. Representative bright field images at time points displayed progressive morphological changes as they differentiated into iMOP-derived neurons (Figure 4B-E). By Day 7, long neurites can be seen extending from cell bodies (Figure 4E arrowheads). To determine the changes in expression levels of Cdkn1b during neuronal differentiation, proliferating iMOP cells and iMOP-derived neurons were compared. Otospheres were labeled with Hoechst and Cdkn1b antibodies. iMOP cells from proliferating otospheres had few cells with low nuclear Cdkn1b expression (Figure 4 F-H). Furthermore, iMOP-derived neurons showed an increase in the number of cells with nuclear Cdkn1b expression 7 days after neuronal differentiation (Figure 4 I-K). To determine if the Cdkn1b cells were adopting a neuronal lineage, labeling of iMOP-derived neurons with the neuronal marker Tubb3 was done. Representative images of iMOP-derived neurons showed co-labeling of Cdkn1b and Tubb3 (Figure 5 A-D). Magnification and quantification of neurites from these cells showed an average 1.5 neurites associated with each Tubb3 labeled cell (n = 60) (Figure 5 E-G). In our neuronal differentiation protocol, immunostaining with Cdkn1b and Tubb3 can be used as an indicator of differentiation into bipolar or pseudo-unipolar iMOP-derived neurons.
Nuclear Cdkn1b Expression Levels Increase after Initiating Differentiation
The morphological changes in otospheres show qualitative changes in iMOP cells as they undergo differentiation. To attain quantitative results from the immunofluorescence images to denote early differentiation events, nuclear fluorescence intensity of Cdkn1b from individual proliferating iMOP cells (n = 239) and iMOP cells undergoing sensory epithelia differentiation (n = 246) were measured. To normalize Cdkn1b fluorescence signals from independent experiments, the ratio of Cdkn1b to Hoechst fluorescence from individual cells was determined. Histograms of the normalized fluorescence intensity of Cdkn1b from proliferating iMOP cells (black) and sensory epithelia differentiating iMOP cells (red) were plotted (Figure 6A). An increase in the number of high Cdkn1b expressing cells was observed as a rightward shift of the histogram for sensory epithelia differentiation (red) relative to the histogram for proliferating iMOP cells (black). To determine the increase in the percentage of cells expressing Cdkn1b, a threshold for normalized Cdkn1b fluorescence intensity units (0.65) was set (arrowhead) and the percentage of cells above the threshold was determined. After sensory epithelia differentiation, iMOP cells displayed an increase from 25.3% to 67.1% of cells expressing Cdkn1b (Figure 6B). The same quantitative analysis was applied to iMOP-derived neurons. When comparing proliferative iMOP cells (n = 525) and 7 day iMOP-derived neurons (n = 583) a rightward shift in the histogram associated with iMOP-derived neurons was observed relative to the histogram for proliferating iMOP cells (Figure 6C). Determining the percentage of cells above the threshold revealed an increase in Cdkn1b expressing cells from 23.5% to 85.5% (Figure 6D). Using the described protocols, quantitative analysis of Cdkn1b expression confirmed an increase in the number of cells that upregulate Cdkn1b during sensory epithelia and neuronal differentiation. The increased numbers of Cdkn1b expressing cells can be used to confirm differentiation of iMOP cells in these protocols.
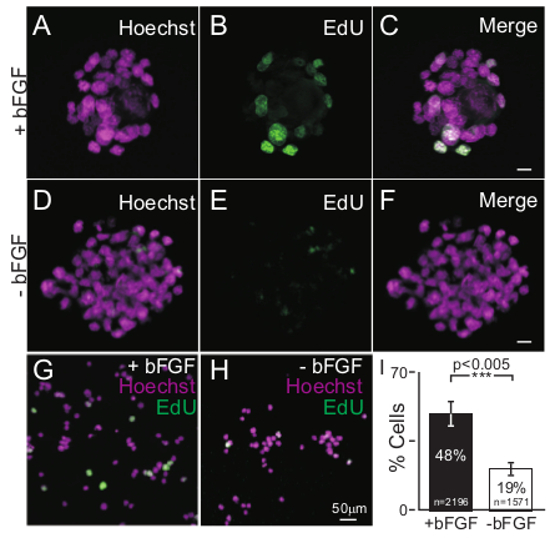
Figure 1. EdU Incorporation as an Assay to Determine Proliferative State of IMOP Cultures. iMOP-derived otospheres cultured in the presence of bFGF. Nuclei of cells were labeled with (A) Hoechst and (B) EdU Alexafluor 488. (C) Merged image of Hoechst and EdU fluorescence. Otospheres cultured 3 days in media lacking bFGF. Cells from cultures were labeled with (D) Hoechst and (E) EdU Alexafluor 488. (F) Merged images of Hoechst and EdU labeling. Merged fluorescence images of Hoechst (magenta) and EdU (green) labeled cells after dissociating otospheres and washing with 1X PBS containing 0.1% Tween 20. Cells were from otospheres cultured (G) in the presence of bFGF or (H) absence of bFGF. Length of scale bars are 10 µm unless indicated.(I) Percent of EdU labeled cells from iMOP cells cultured in the presence or absence of bFGF. The number of individual cells analyzed was denoted within the bar graph and error bars were depicted as standard error of the mean (SEM). Cell counts were from n = 5 independent experiments. The Student's t-test was done to determine statistical significance. Please click here to view a larger version of this figure.
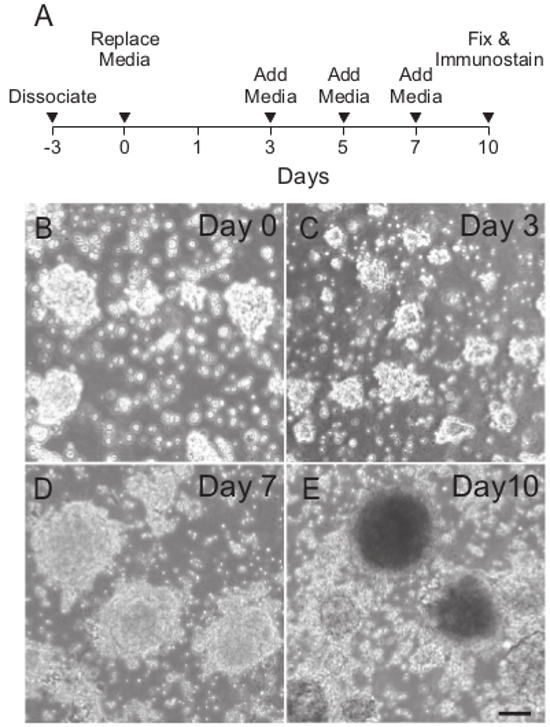
Figure 2. General Schematic of Sensory Epithelia Differentiation. (A) Timeline for differentiating iMOP cells into sensory epithelia. The line graph denotes the time of cell dissociation and media changes. (B) Typical phase contrast images of otospheres on Day 0, (C) 3, (D) 7 and (E) 10 during unguided sensory epithelia differentiation. Length of scale bar is 100 µm. Please click here to view a larger version of this figure.
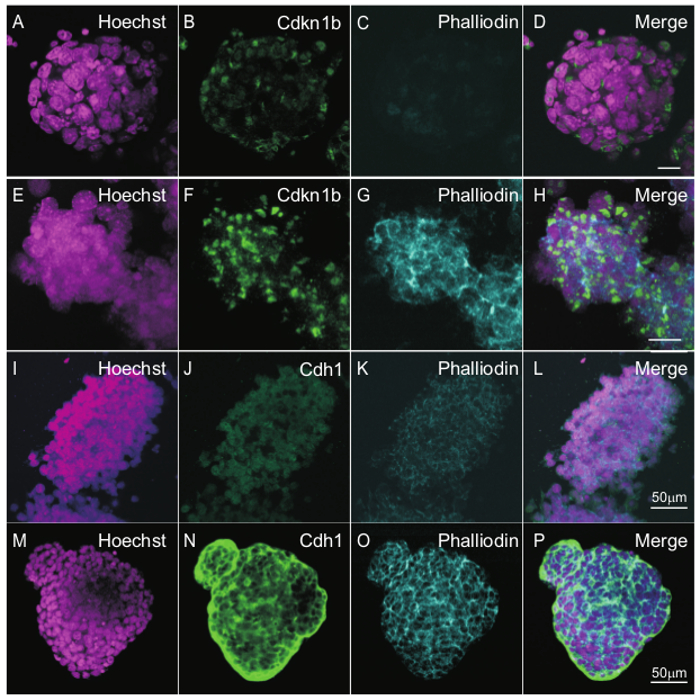
Figure 3. Expression of Markers Indicative of Cell Cycle Arrest and Differentiation in IMOP-derived Sensory Epithelia. Otosphere of iMOP cells cultured in iMOP culture media. Nuclei of cells were labeled with (A) Hoechst and (B) Cdkn1b antibody. Filamentous actin was labeled with (C) phalloidin. (D) The merged image of a typical otospheres containing proliferating iMOP cells. Otospheres from iMOP cells were cultured in sensory epithelia culture media for 10 days. Cells from otopsheres were marked with (E) Hoechst, (F) Cdkn1b and (G) phalloidin. (H) Merged image of otospheres showing increased Cdkn1b and phalloidin labeling. Proliferating otospheres were labeled with (I) Hoechst, (J) Cdh1, (K) phalloidin. (L) The merged image shows weak fluorescence from these markers. Otospheres differentiated into sensory epithelia were also labeled with (M) Hoechst, (N) Cdh1, (O) phalloidin. (P) The merged image shows typical strong Cdh1 and phalloidin labeling. Length of scale bars are 10 µm unless indicated. Please click here to view a larger version of this figure.
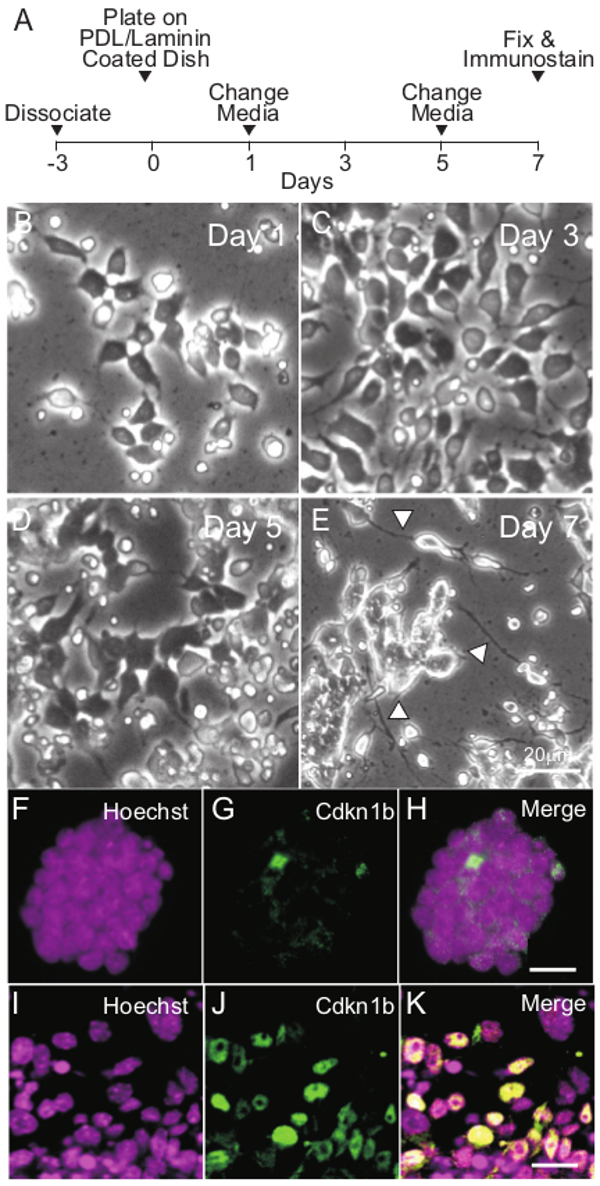
Figure 4. General Schematic of Neuronal Differentiation. (A) Timeline for differentiating iMOP cells into neurons. Phase contrast images of iMOP cells undergoing neuronal differentiation at (B) Day 1, (C) 3, (D) 5 and (E) 7. Arrowheads point to neurites. Fluorescence images of iMOP cells cultured as otospheres in iMOP culture media labeled with (F) Hoechst and (G) Cdkn1b. (H) Merged image of cells labeled with Hoechst and Cdkn1b. Fluorescence images of iMOP cells after 7 days of culture in neuronal differentiation media. Nuclei of cells were labeled with (I) Hoechst and (J) Cdkn1b. (K) Merged image of cells labeled with Hoechst and Cdkn1b. Length of scale bars are 10 µm unless noted. Please click here to view a larger version of this figure.
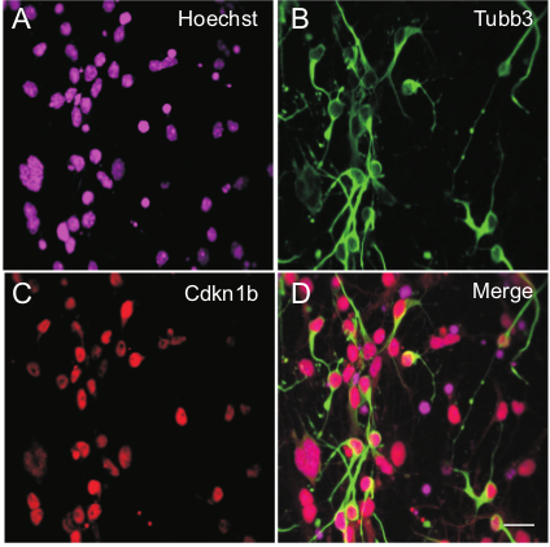
Figure 5. Expression of Molecular Markers Indicative of Cell Cycle Exit and Neuronal Differentiation. iMOP cells 7 days after neuronal differentiation. Nuclei were labeled with (A) Hoechst, (B) Tubb3 (neuronal β-tubulin) antibodies to highlight cellular morphology and (C) Cdkn1b antibodies. (D) Merged image with labeling of Cdkn1b and Tubb3 in individual cells. Length of scale bar is 20 µm. Please click here to view a larger version of this figure.
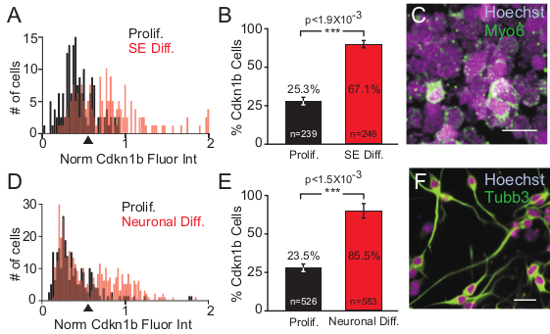
Figure 6. Single Cell Quantitative Fluorescence Intensity Analysis of Cdkn1b. (A) Normalized fluorescence intensity of Cdkn1b expression was determined by calculating the ratio of Cdkn1b and Hoechst fluorescence intensity from individual cells. The normalized fluorescence intensity was plotted relative to cell numbers as a histogram. Normalized fluorescence intensity of Cdkn1b from proliferating iMOP cells cultured in the presence of bFGF (black) and iMOP otospheres differentiated into sensory epithelia (SE) (red) are shown. A threshold was set at 0.65 normalized fluorescence intensity units (arrowhead) (B) Percentage of iMOP cells expressing Cdkn1b above the threshold value, from proliferating (black) and sensory epithelia differentiated cultures (red). (C) Merged fluorescent image of sensory epithelia differentiated iMOP cells labeled with Hoechst and Myo6 antibodies. (D) Normalized fluorescence intensity of Cdkn1b expression from proliferating iMOP cells (black) and neuronal differentiated iMOP cells (red) were plotted and a threshold set (arrowhead) as described above. (E) Percentage of iMOP cells expressing Cdkn1b from proliferating (black) and neuronal differentiating cultures (red). Individual cells used for analysis were denoted in each bar graph. Results were compiled from different experiments (n = 3) and error bars depicted as SEM. The Student t-test was used to determine the statistical significance. (F) Merged fluorescent image of bipolar or pseudo-unipolar iMOP-derived neurons labeled with Hoechst and Tubb3. Length of scale bars are 20 µm. Please click here to view a larger version of this figure.
| Cell Culture Vessel | Surface Area (cm2) | Number of Cells Seeded for Routine Maintenance | Number of Cells Seeded for Sensory Epithelia Differentiation | Number of Cells Seeded for Neuronal Differentiation | Scaling Factor Relative to 60 mm Dish | Volume of Media Used (ml) | |
| Multwell Plates | |||||||
| 96 | 0.3 | 10,000 | 20,000 | 10,000-50,000 | 0.015 | 0.1 | |
| 24 | 2 | 90,000 | 180,000 | 100,000-150,000 | 0.100 | 0.5 | |
| 12 | 4 | 180,000 | 360,000 | 300,000 | 0.200 | 1 | |
| 6 | 10 | 500,000 | 1,000,000 | 500,000-750,000 | 0.500 | 2 | |
| Dishes | |||||||
| 35 mm | 10 | 500,000 | 1,000,000 | 500,000 | 0.500 | 2 | |
| 60 mm | 20 | 1,000,000 | 2,000,000 | 1,000,000-1,500,000 | 1.000 | 3 | |
| 100 mm | 60 | 2,500,000 | 5,000,000 | 2,500,000- 3,000,000 | 3.000 | 8 | |
Table 1. Cell Numbers for Maintenance of Differentiation of IMOP Cells in Various Tissue Culture Plate Formats.
