A Whole Mount In Situ Hybridization Method for the Gastropod Mollusc Lymnaea stagnalis
Summary
The goal of this protocol is to provide users with a set of methods for the high-throughput decapsulation of Lymnaea stagnalis embryos and larvae in preparation for whole mount in situ hybridization, and for subsequent pre- and post-hybridization treatments.
Abstract
Whole mount in situ hybridization (WMISH) is a technique that allows for the spatial resolution of nucleic acid molecules (often mRNAs) within a 'whole mount' tissue preparation, or developmental stage (such as an embryo or larva) of interest. WMISH is extremely powerful because it can significantly contribute to the functional characterization of complex metazoan genomes, a challenge that is becoming more of a bottleneck with the deluge of next generation sequence data. Despite the conceptual simplicity of the technique much time is often needed to optimize the various parameters inherent to WMISH experiments for novel model systems; subtle differences in the cellular and biochemical properties between tissue types and developmental stages mean that a single WMISH method may not be appropriate for all situations. We have developed a set of WMISH methods for the re-emerging gastropod model Lymnaea stagnalis that generate consistent and clear WMISH signals for a range of genes, and across all developmental stages. These methods include the assignment of larvae of unknown chronological age to an ontogenetic window, the efficient removal of embryos and larvae from their egg capsules, the application of an appropriate Proteinase-K treatment for each ontogenetic window, and hybridization, post-hybridization and immunodetection steps. These methods provide a foundation from which the resulting signal for a given RNA transcript can be further refined with probe specific adjustments (primarily probe concentration and hybridization temperature).
Introduction
Molluscs are a group of animals that hold the interest of a broad diversity of scientific disciplines. Despite their morphological diversity1, species richness (second only to the Arthropods in terms of species number2) and relevance to a wide range of commercial3, medical4 and scientific issues5-8, there are relatively few molluscan species that can claim to be both well-equipped scientific models and easy to maintain in a laboratory environment. One mollusc that is much used by disciplines such as neurobiology9, ecotoxicology10 and more recently evolutionary biology11,12, is Lymnaea stagnalis, primarily because of its widespread distribution and extreme ease of maintenance. Despite its popularity as a 'model' organism and its long history of use by developmental biologists13-19, the range and power of molecular tools available to the L. stagnalis scientific community lies far behind that of more traditional animal models (Drosophila, mouse, sea urchin, nematodes).
Our desire to develop Lymnaea as a molecular model stems from an interest in the molecular mechanisms that guide shell formation. This motivated us to refine a set of techniques that would allow for the efficient, consistent and sensitive visualization of gene expression during Lymnaea's development. Whole mount in situ hybridization (WMISH) is widely employed for a variety of model organisms and has been in use for more than 40 years 20. In its different guises, ISH can be employed to spatially localize specific loci on chromosomes, rRNA, mRNA and micro-RNAs.
One of the challenges we needed to address prior to refining a WMISH method for L. stagnalis was the issue of gently and efficiently extracting embryos and larvae of varying stages from the egg capsules in which they are deposited. This extraction, or 'decapsulation', needs to be achieved efficiently in order to collect adequate material for a given in situ experiment, while at the same time maintaining morphological and cellular integrity. While other model organisms also undergo encapsulated development, in our hands none of the methods reported for those species could be successfully employed in L. stagnalis.
The overall goals of this method are therefore: to extract L. stagnalis embryos and larvae from their capsules in a high-throughput fashion, to apply pre-hybridization treatments that optimize the WMISH signal, to prepare embryos and larvae with satisfactory WMISHsignals for imaging.
Protocol
NOTE: The following steps outline our method for conducting an in situ experiment on embryonic and larval stages of L. stagnalis. Where a step involves the use of a hazardous chemical this is indicated by the word 'CAUTION' and all appropriate safety procedures should be adopted. Links to representative MSDS sheets for hazardous chemicals are provided in Supplementary File 1. Recipes for all reagents are provided in Supplementary File 2.
1. Assembly of Decapsulation Apparatus
- To do this, connect a 20 ml disposable syringe to silicon tubing (with an inner diameter of 1 mm and an outer diameter of 3 mm) using a P1,000 tip cut to an appropriate length as shown in Figure 2A.
- Tape a standard microscope slide to an inverted large Petri dish. Tape the silicon tubing immediately adjacent to the microscope slide as shown in Figure 2C and 2D.
- Rest a pulled glass needle on the microscope slide and gently insert it into the silicon tubing until the tip of the needle protrudes approximately 20% of the way across the inner diameter of the tubing (see Figure 2C and 2D). Once the needle is in position tape it down to the Petri dish.
- Allow the free end of the silicon tubing to rest in another Petri dish that will collect the decapsulated material.
2. Sample Collection, Fixation and Decapsulation
NOTE: All steps are carried out at RT unless otherwise noted.
- Carefully collect egg strings from the walls of an aquarium. To do this, use a flat piece of flexible plastic as a spatula to scrape the egg string off the substrate, and use a plastic tea strainer to fish the floating egg string out of the water. Stage and sort the material under a microscope using the guide provided in Figure 1.
- Place the egg string onto a paper towel and make a longitudinal incision along the egg mass using the featherweight forceps. Roll the egg capsules out of the egg string and remove as much of the jelly material as possible from each capsule by pushing them around the paper towel using the featherweight forceps.
- Using the featherweight forceps, transfer the egg capsules into a Petri dish containing 5 ml of tap water. Continue to collect de-jellied egg capsules of the desired developmental stages into this dish. Collect enough capsules from all developmental stages for the planned WMISH experiment then proceed to the next step.
- When working with more developed larvae (5 days post first cleavage (dpfc) and older) anaesthetize them prior to fixation.
NOTE: This will prevent muscles from contracting which makes the interpretation of in situ staining patterns very difficult.- Relax larvae (while they are still in their capsules) in a 2% w/v solution of MgCl2•6H2O for 30 min prior to fixation.
- Assess the degree of relaxation after 30 min by submerging several larvae while still in their egg capsules in fixative solution and monitoring their response under a microscope. Incompletely relaxed larvae will retract into their shells, while fully relaxed larvae will not respond. Once these larvae have been relaxed proceed to the next step.
- Transfer the egg capsules using a wide bore plastic pipette into a sealable tube that provides 10 times the volume of egg capsules (e.g., 1 ml of settled capsules would require a 10+ ml tube).
- Aspirate as much liquid as possible from the tube and replace with a volume of fixative solution that is 10x the volume of the settled egg capsules. Gently rotate the egg capsules in fixative at RT for the appropriate time for each developmental stage (see Figure 1).
- Discontinue rotation and allow the capsules to sink and aspirate the fixative solution into an appropriate waste container.
- Wash the egg capsules by replacing the fixative solution with Phosphate Buffered Saline with 0.1% Tween-20 (PBTw) and rotating at RT for 5 min. Aspirate the PBTw and repeat twice.
- Remove embryos and larvae from their capsules using the apparatus (see section 1 for details) shown in Figure 2. Draw the capsules up into the 20 ml syringe, attach the tubing and then dispel the capsules through the tubing and past the glass needle out into the collection dish.
NOTE: Normally, the majority (>90%) of all capsules should need only one pass through the device. In many cases the capsule membrane is damaged but embryos and larvae remain inside the ruptured capsule. Reprocess this material by simply drawing it up into the syringe and dispelling it again. - Collect the decapsulated embryos and larvae into a 1.5 ml tube using a micropipette (a P20 for 0 – 3 day old larvae, and a P200 for older larvae with the end of the tip cut off).
NOTE: A pause (for up to several months) in the protocol can be made at this stage. If this is required, the decapsuled material should be collected into a 1.5 ml tube for storage (continue to the next step). Otherwise continue immediately to Protocol 3. - Allow embryos and larvae to sink to the bottom of the tube and aspirate the supernatant. Replace with 33% Ethanol (EtOH) in PBTw and let sit for 5 – 10 min. Repeat with 66% EtOH in PBTw and 100% EtOH. Wash larvae twice in 100% EtOH at RT. Store the material at -20 °C in 100% EtOH.
- When ready to continue, re-hydrate the samples by removing the 100% EtOH and replacing with 66% EtOH in PBTw, let sit for 5 – 10 min. Repeat with 33% EtOH in PBTw, let sit for 5 – 10 min. Finally wash with 3x 5 min washes of PBTw to ensure all EtOH is removed.
3. Proteinase-K, TEA and Post-fixation
NOTE: We find performing the following steps in small baskets with a mesh floor the most efficient and gentle method for quickly exchanging time critical solutions. While these can be home made, we use baskets (medium size) that are compatible with the Intavis InSituPro-Vsi liquid handling robot (www.intavis.de/products/automated-ish-and-ihc). Such baskets can be quickly and easily moved between the wells of a 12 well tissue culture dish (TCD) in order to exchange solutions, or the solution can be aspirated from the well using a pipette. Alternatively, all solution exchanges can be performed without these baskets by simply aspirating the supernatant from the larvae. In this case a gentle swirling motion will concentrate all embryos and larvae to the center of the dish allowing the supernatant to be removed from the edge of the well. The following assumes the user is employing baskets for solution exchanges.
- Using a pipette, transfer embryos and larvae into a basket sitting in a 12 well TCD with 2 ml of PBTw.
NOTE: The number of embryos and/or larvae that can be added depends on the developmental stage being investigated, however a general rule of thumb is to maintain at least 25% of the floor space free of embryos/larvae (i.e., do not overcrowd the basket). - Prepare an adjacent well with 2 ml of the appropriate Proteinase-K solution (see Figure 1), and another 2 wells with 2 ml of 0.2% Glycine each. Move each basket into the well containing the appropriate concentration of Proteinase-K and immediately start timing.
- After 10 min move each basket into a well containing 0.2% Glycine and incubate for 5 min. Then move each basket into the second well with 0.2% Glycine and incubate for 5 min. Wash out the Glycine with 3x 5 min exchanges of 3 ml PBTw.
- Remove the PBTw solution and treat the samples once with freshly prepared Triethanolamine (TEA) solution for 5 min CAUTION! Do not agitate. Replace with 3 ml of freshly prepared TEA solution for 5 min. Do not agitate. Aspirate the majority of the TEA solution from the samples. Add the freshly prepared Triethanolamine with Acetic Anhydride (TEAAA) solution and incubate for 5 min. CAUTION! Do not agitate.
- OPTIONAL – While the above step is incubating, prepare another fresh batch of TEAAA solution and repeat the above step.
NOTE: This second treatment with TEAAA is optional but may help to completely eliminate all background with probes prone to generating background. - Remove the TEAAA solution by aspirating it from the well and replace with 3 ml of PBTw. Do not agitate. Remove the PBTw and apply 3 ml of 3.7% formaldehyde in PBTw. Gently swirl occasionally during a 30 min incubation.
- Remove the fixative by aspirating it from the well and replace with 3 ml of PBTw. Transfer the material into a 1.5 ml tube. Replace the PBTw with hybridization buffer and incubate for 5 min at RT – CAUTION.
- Place the tubes into a hot block at RT, and set the temperature to the desired hybridization temperature.
NOTE: The hybridization temperature is probe specific and will need to be empirically optimized, however we find 55 °C to be a good temperature to initially trial. Allow the block to come to the hybridization temperature, and allow the samples to pre-hybridize for approximately 15 min (i.e., the time it takes to prepare the riboprobes, longer is not necessary). Prepare adequate volumes (normally 100 µl) of the diluted riboprobes. We typically trial probe concentrations of 100 and 500 ng/ml as an initial range. We prepare riboprobes according to 12. - Remove the hybridization buffer from the samples and add the probe in hybridization buffer to the samples. Overlay with 100 µl (or an adequate volume to form a phase above the hybridization buffer) of mineral oil.
NOTE: The mineral oil prevents extensive condensation that will form during the lengthy hybridization step. Extensive condensation will significantly alter both the chemistry of the hybridization solution and the concentration of the probe. - Denature the probe and the target RNA by heating the samples to 75 °C for 10 min, then reduce the heat to the desired hybridization temperature. Allow hybridization to proceed for a minimum of 12 hr (O/N) or longer (24 – 48 hr).
- During hybridization remove a single tube from the heat block, rotate it rapidly between thumb and forefinger to suspend the larvae without disturbing the oil phase, and replace it in the heat block. Repeat this once every 6 – 12 hr or so.
4. Hot Washes and Immunodetection
NOTE: While we use a liquid handling robot for the following steps, these can also easily be done manually. In this case, embryos and larvae should be kept in the 1.5 ml tubes they were hybridized in. All subsequent solution exchanges are aspirated and added with a P1,000 pipette. When performed manually each of the following steps should employ 1 ml of each solution.
- Heat adequate volumes (3 ml each for each sample) of the 4x, 2x and 1x wash solutions to the hybridization temperature.
- Wash all samples three times in 4x wash buffer for 15 min each at the hybridization temperature. Wash all samples three times in 2x wash buffer for 15 min each at the hybridization temperature. Wash all samples three times in 1x wash buffer for 15 min each at the hybridization temperature.
- Wash all samples once with 1x Sodium Chloride Sodium Citrate buffer + 0.1% Tween (SSC + 0.1% Tween) at the hybridization temperature. Allow samples to cool to RT.
- Wash all samples twice in 1x SSC + 0.1% Tween for 15 min Replace this 1x SSC solution with Maleic Acid Buffer (MAB) and let sit for 10 min Repeat the MAB wash. Replace MAB with block solution and incubate for 1.5 hr.
- Exchange the block solution for antibody solution (1:10,000 dilution of antibody in block solution) and incubate for 12 hr (O/N) at RT with gentle agitation.
5. Color Development and Mounting
- Aspirate the antibody solution and wash 15 times with PBTw for 10 min each. Replace PBTw with 1x Alkaline Phosphatase Buffer (AP) and incubate for 10 min.
- While the above incubation step proceeds, prepare the AP staining solution (see Supplementary File 2) – CAUTION. Transfer the material into a TCD well and replace the 1x AP solution with AP staining solution. Note the time now so that the length of time the color reaction takes place for can be recorded. Monitor the development of the staining pattern until the signal to background ratio is optimal.
- To stop color development, remove the 1x AP staining solution (dispose in the appropriate waste container), and apply 2 ml of PBTw and note the time now. Rinse twice more with PBTw for 5 min each. Use one of these rinses to transfer the material into a 1.5 ml tube.
- Remove the PBTw and apply 1 ml of 3.7% formaldehyde in PBTw and agitate or rotate for least 30 min at RT (this can also be done O/N).
- Wash out the fixative with 3 washes of PBT (dispose the waste fixative in an appropriate waste container). Wash the samples 3 times at 50 °C in de-ionized water.
NOTE: These washes eliminate the precipitation of salts that may become visible during clearing and visualization steps. - Decide whether the samples should be mounted in Benzyl Benzoate:Benzyl Alcohol (BB:BA, also known as Murray's clear) or glycerol.
NOTE: We prefer the more powerful clearing agent BB:BA as L. stagnalis embryos are somewhat opaque. Samples to be mounted in BB:BA will first need to be dehydrated through an ethanol series. Samples to be mounted in 60% glycerol can be immediately mounted. - Transfer the samples into the mounting solution (BB:BA or 60% glycerol) using a pipette.
NOTE: When mounting in BB:BA this must be done in a glass well (BB:BA will melt polycarbonate plastic). - Allow the samples to clear for 5 – 10 min, and then mount them onto a slide using an appropriate number of stacked coverslips as spacers (1 coverslip for samples <1 dpfc, 2 coverslips for samples >1 dpfc).
NOTE: Whole mounts can now be imaged using a compound microscope with DIC optics.
Representative Results
The representative WMISH staining patterns shown in Figure 3 were generated using the technique described above, and reflect a variety of spatial expression patterns for genes involved in a range of molecular processes ranging from shell formation (Novel gene 1, 2, 3 and 4), to cell-cell signaling (Dpp) to transcription regulation (Brachyury) across a range of developmental stages. While we have not quantified the expression levels of these genes we expect that they would also vary significantly, indicating that our method can be applied against a broad variety of gene products expressed in all stages of development at various levels. Only one of the genes presented here (Dpp) has been previously described in L. stagnalis 21,22. The results we present here are largely in keeping with these previous reports, but with significantly higher spatial resolution. The spatial expression pattern of Brachyury has been described in abalone 23 and limpet 24 and in both cases was also detected in mantle cells as we find for L. stagnalis (Figure 3F). We isolated Novel genes 1 – 4 (from a proteomic screen designed to identify gene products directly involved in shell formation, and so their spatial expression patterns associated with the shell gland (Figure 3A and B) or shell field (Figure 3C and D) are completely congruent with shell-forming functions. These results indicate that the high throughput technique we have developed for removing embryos and larvae from the egg capsule, and the subsequent stage-specific permeabilization treatments, generate whole mount samples that will yield high quality in situ staining patterns for a wide variety of genes for all stages of embryonic and larval development.
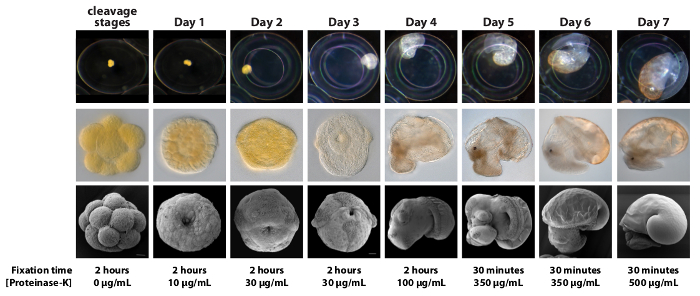
Figure 1. A Summarized Ontogeny of Lymnaea stagnalis and Corresponding Fixation and Proteinase-K Treatments. Representative images of embryos and larvae from the first 7 days of development illustrate a significant increase in size (row 1) and morphological complexity (rows 2 and 3). These developmental changes translate into large differences in the appropriate fixation time and Proteinase-K concentrations that generate optimal WMISH signals. For WMISH all stages should be fixed in 3.7% formaldehyde in PBS at RT with gentle agitation within their egg capsules. All stages should then be treated with the appropriate Proteinase-K concentration for 10 min. Note that we observe significant inter-batch variation in the activity of Proteinase-K from our supplier. This variation must be accounted for by performing a round of 'calibrating' WMISH experiments where the activity of the new Proteinase-K is empirically determined. The Proteinase-K concentrations stated in the figure should therefore be treated as an initial guide, however the relative concentrations between developmental stages (for example 2 day old embryos require a Proteinase-K concentration 3 times higher than day 1 embryos) are set. Please click here to view a larger version of this figure.
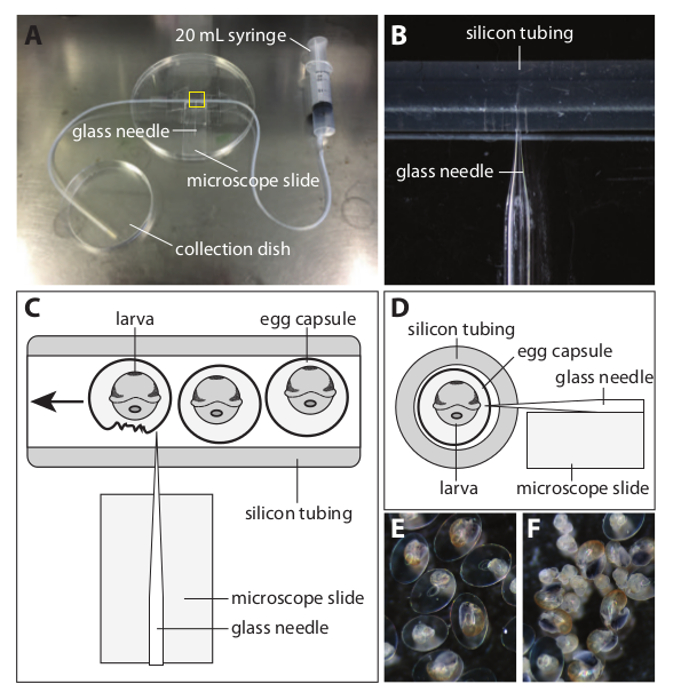
Figure 2. Apparatus Used to Decapsule L. stagnalis Embryos. (A) An overview of the apparatus that can efficiently remove L. stagnalis embryos and larvae from their capsules. (B) A magnified view of the yellow boxed area in (A). A sharp glass needle is placed on the microscope slide and inserted into the silicon tubing (inner diameter 1 mm, outer diameter 3 mm) such that the tip protrudes approximately 20% of the way into the cavity of the tubing. The glass needle is then also taped to the microscope slide and the Petri dish. (C) A schematic 'plan' view of the yellow boxed section in A. Egg capsules containing fixed embryos and larvae are first collected using the 20 ml syringe. The syringe is then attached to the silicon tubing and the capsules expelled through the tubing and past the needle. Egg capsule membranes are torn by the needle and the liberated embryonic and larval material can be collected from the collection dish using a micropipette. (D) A schematic 'cross section' view of the yellow boxed section in A. The microscope slide ensures that the needle enters the silicon tubing at the correct height. (E) A representative view of larvae prior to being processed by the decapsuling apparatus. (F) A representative view of larvae that have made a single pass through the apparatus. More than 90% of the material has been effectively and gently removed from their capsules. Please click here to view a larger version of this figure.
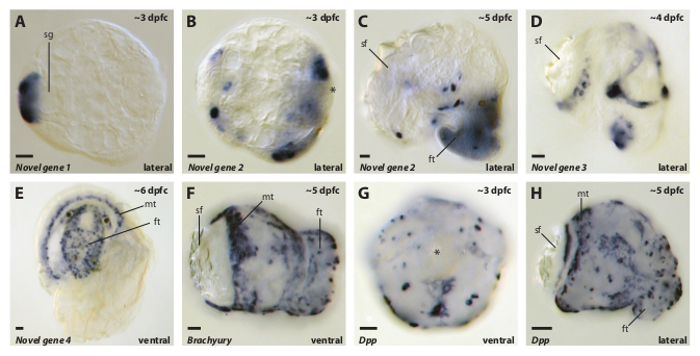
Figure 3. Representative Images of WMISH Expression Patterns Against a Variety of Genes From a Range of L. stagnalis Developmental Stages Generated by the Method Described Here. All developmental stages were processed as described in the above method and have been mounted and imaged in BB:BA (Murray's clear). Approximate ages are indicated in the top right of each panel and the orientation is indicated in the lower right. Gene orthology (when known) is indicated in the lower left of each panel. Abbreviations: shell gland (sg); shell field (sf); mantle (mt); foot (ft); Decapentaplegic (Dpp); dpfc (days post first cleavage). All scale bars are 20 µm. Please click here to view a larger version of this figure.
Discussion
The method described here allows for the efficient visualization of RNA transcripts with presumably varying expression levels within all developmental stages of Lymnaea stagnalis. To remove embryos and larvae from their capsules we trialed a variety of chemical, osmotic shock and physical treatments reported for other encapsulated-developing model organisms. However, in our hands the method we describe here is the only high-throughput technique that removes the tough capsular membrane without damaging the embryos and larvae. Following decapsulation, the material can either be stored, or treated with a stage specific regimen of Proteinase-K and then hybridized to a riboprobe. Additional empirical optimization efforts (typically focused on probe concentration and hybridization temperature) may be required for each probe/target. These parameters (in addition to the fixation regimen and Proteinase-K treatments) are typically the most influential parameters of any in situ experiment (assuming that the quality of the fixed material and the RNA probe are of a high standard).
The importance of an appropriate Proteinase-K treatment to the final result of an in situ experiment is paramount for L. stagnalis. This is reflected in the wide range of Proteinase-K concentrations required by distinct developmental stages (ranging from 0 µg/ml to 500 µg/ml). It is therefore important to be able to assign a given egg string to an ontogenetic window. To this end, the guideline that we provide in Figure 1 allows for the staging of developmental material of unknown ages (a print friendly version of this figure is available in Supplementary File 3 that users may find useful to have at the microscope when staging material). We note that for other species of gastropods Proteinase-K treatments for WMISH can either be kept constant for a wide range of developmental stages 8,25,26, or can be omitted entirely 27. This is in stark contrast to the situation in L. stagnalis. Furthermore, while other research groups have previously reported WMISH expression patterns for several genes in L. stagnalis larvae (see 22,28,29) the method that we describe here yields patterns of significantly higher spatial resolution. Finally, we have observed significant inter-batch variation in the activity of the Proteinase-K from our supplier. This variation must be accounted for by performing a round of 'calibrating' WMISH experiments where the activity of the new Proteinase-K is empirically determined. All subsequent experiments with aliquots of Proteinase-K from that batch can then be freely performed.
We previously described an alternative WMISH method for L. stagnalis embryos and larvae elsewhere 12. That method detailed the use of the mucolytic agent N-acetyl-L-cysteine (NAC), a reducing agent such as Dithiothreitol (DTT) and a pre-hybridization treatment with sodium dodecyl sulfate (SDS). We found those treatments enhanced the staining patterns of some genes for some developmental stages. The fixation strategy that we recently developed and describe here (fixing larvae within their capsules) simplifies and expedites the steps required to prepare material for an in situ experiment, and apparently negates the need for empirically determining additional optimal pre-hybridization treatments with NAC, DTT or SDS. Future refinements to the technique reported here could include the visualization of microRNAs (following modifications to standard WMISH protocols previously reported 30), double or triple labeling of mRNA targets 31, and fluorescent visualization of WMISH signals 32. Arguably the greatest limitation of the technique is the overall length of time it takes to go from collecting the material, to a digital image that represents a given gene expression pattern. Due to the nature of the biochemical and biophysical events that must take place during such a process this is an inherent feature of most in situ hybridization protocols.
Lymnaea occupies a position within the Metazoa that is extremely under-represented in terms of model organisms. As a representative Spiralian, Lymnaea can bring insight into the evolution of distinct morphological features such as shell formation 12 and body handedness 33-35 and is also a valuable neuroethology 36 and neurophysiology model 9,37. Powerful techniques such as the ability to efficiently visualize gene expression patterns in situ increases the functionality of Lymnaea as a model organism, and broadens the variety of questions that it can be used to address. At a time when the generation of large sequence datasets (complete transcriptomes and even genomes) is relatively routine, such methods will become more relevant to researchers wishing to interpret the flood of sequence data from such models. While Lymnaea is a relatively derived gastropod 38, and possesses what would be considered a large genome in comparison to other model organisms (1.22 Gb 39), it has many practical and interesting features that make it an attractive model system. The methods that we describe here expand the toolbox available to Lymnaea and may be of use to other species that undergo encapsulated development.
Disclosures
The authors have nothing to disclose.
Acknowledgements
This work was supported by funding to DJJ through DFG project #JA2108/2-1.
Materials
| Featherweight forceps | Ehlert & Partner | #4181119 | |
| Silicon tubing | Glasgerätebau OCHS GmbH | 760070 | |
| Glass capillaries | Hilgenberg | 1403547 | |
| 12 well tissue culture dishes | Carl Roth | CE55.1 | |
| 37% Formaldehyde | Carl Roth | P733.1 | CAUTION – May cause cancer. Toxic by inhalation, in contact with skin and if swallowed. Toxic: danger of very serious irreversible effects through inhalation, in contact with skin and if swallowed. |
| Ethylenediamine tetraacetic acid | Carl Roth | CN06.3 | CAUTION – CAUSES EYE IRRITATION. MAY CAUSE RESPIRATORY TRACT AND SKIN IRRITATION. Avoid breathing dust. Avoid contact with eyes, skin and clothing. Use only with adequate ventilation |
| Magnesium Chloride | Carl Roth | 2189.1 | |
| Tween-20 | Carl Roth | 9127.1 | CAUTION – May be harmful if inhaled. May cause respiratory tract irritation. May be harmful if absorbed through skin. May cause skin irritation. May cause eye irritation. May be harmful if swallowed. |
| Sodium Chloride | Carl Roth | 3957.1 | |
| Ficoll type 400 | Carl Roth | CN90.1 | |
| polyvinylpyrrolidone K30 (MW 40) | Carl Roth | 4607.1 | CAUTION – May be harmful if inhaled. May cause respiratory tract irritation. May be harmful if absorbed through skin. May cause skin irritation. May cause eye irritation. May be harmful if swallowed. |
| Nuclease freeBovine Serum Albumin | Carl Roth | 8895.1 | |
| Salmon sperm | Carl Roth | 5434.2 | |
| Heparin | Carl Roth | 7692.1 | CAUTION – ADVERSE EFFECTS INCLUDE HEMORRHAGE, LOCAL IRRITATION. POSSIBLE ALLERGIC REACTION IF INHALED, INGESTED/CONTACTED. EYES/SKIN/RESPIRATORY TRACT IRRITANT. POSSIBLE HYPERSENSITIZATION. DURING PREGNANCY HAS BEEN REPORTED TO INCREASE RISK OF STILLBIRTH |
| Proteinase-K | Carl Roth | 7528.1 | |
| Glycine | Carl Roth | 3790.2 | |
| Deionised formamide | Carl Roth | P040.1 | CAUTION – Irritating to eyes and skin. May be harmful by inhalation, in contact with skin and if swallowed. May cause harm to the unborn child. Hygroscopic. |
| Standard formamide | Carl Roth | 6749.3 | CAUTION – Irritating to eyes and skin. May be harmful by inhalation, in contact with skin and if swallowed. May cause harm to the unborn child. Hygroscopic. |
| Triethanolamine | Carl Roth | 6300.1 | CAUTION – Avoid breathing vapor or mist. Avoid contact with eyes. Avoid prolonged or repeated contact with skin. Wash thoroughly after handling. |
| Acetic anhydride | Carl Roth | 4483.1 | CAUTION – CAUSES SEVERE SKIN AND EYE BURNS. REACTS VIOLENTLY WITH WATER. HARMFUL IF SWALLOWED. VAPOR IRRITATING TO THE EYES AND RESPIRATORY TRACT |
| Maleic acid | Carl Roth | K304.2 | CAUTION – Very hazardous in case of eye contact (irritant), of ingestion, . Hazardous in case of skin contact (irritant), of inhalation (lung irritant). Slightly hazardous in case of skin contact (permeator). Corrosive to eyes and skin. |
| Benzyl benzoate | Sigma | B6630-250ML | CAUTION – May be harmful if inhaled. May cause respiratory tract irritation. May be harmful if absorbed through skin. May cause skin irritation. May cause eye irritation. Harmful if swallowed. |
| Benzyl alcohol | Sigma | 10,800-6 | CAUTION – Harmful if swallowed. Harmful if inhaled. Causes serious eye irritation. |
| Glycerol | Carl Roth | 3783.1 | |
| Blocking powder | Roche | 11096176001 | |
| Anti DIG Fab fragments AP conjugated | Roche | 11093274910 | |
| Tris-HCl | Carl Roth | 9090.3 | |
| 4-Nitro blue tetrazolium chloride in dimethylformamide | Carl Roth | 4421.3 | CAUTION – May cause harm to the unborn child. Harmful by inhalation and in contact with skin. Irritating to eyes. |
| 5-bromo-4-chloro-3-indolyl-phosphate | Carl Roth | A155.3 | CAUTION – Potentially harmful if ingested. Do not get on skin, in eyes, or on clothing. Potential skin and eye irritant. |
| N-acetyl cysteine | Carl Roth | 4126.1 | |
| Dithiothreitol | Carl Roth | 6908.1 | CAUTION – May cause eye and skin irritation. May cause respiratory and digestive tract irritation. The toxicological properties of this material have not been fully investigated. |
| Tergitol | Sigma | NP40S | CAUTION – May be harmful if inhaled. May cause respiratory tract irritation. May be harmful if absorbed through skin. May cause skin irritation. May cause eye irritation. May be harmful if swallowed. |
| Sodium dodecyl sulphate | Carl Roth | CN30.3 | CAUTION – Harmful if swallowed. Toxic in contact with skin. Causes skin irritation. Causes serious eye damage. May cause respiratory irritation. |
| Potassium Chloride | Carl Roth | 6781.1 | |
| di-Sodium hydrogen phosphate dihydrate (Na2HPO4.2H2O) | Carl Roth | 4984.1 | |
| Potassium dihydrogen phosphate (KH2PO4) | Carl Roth | 3904.1 | |
| Tri sodium citrate dihydrate (C6H5Na3O7.2H2O) | Carl Roth | 3580.1 | CAUTION – May cause eye, skin, and respiratory tract irritation. The toxicological properties of this material have not been fully investigated. |
| Mineral oil | Carl Roth | HP50.2 | |
| InSituPro-Vsi | Intavis | www.intavis.de/products/automated-ish-and-ihc |
References
- Smith, S. A., Wilson, N. G., Goetz, F. E., Feehery, C., Andrade, S. C. S., et al. Resolving the evolutionary relationships of molluscs with phylogenomic tools. Nature. 480 (7377), 364-367 (2011).
- Brusca, R. C., Brusca, G. J. . Invertebrates. , (2002).
- World Health Organization. Schistosomiasis: number of people treated in 2011. Week. Epi. Rec. 88, 81-88 (2013).
- Henry, J. Q., Collin, R., Perry, K. J. The slipper snail, Crepidula.: an emerging lophotrochozoan model system. Biol. Bull. 218 (3), 211-229 (2010).
- Perry, K. J., Henry, J. Q. CRISPR/Cas9-mediated genome modification in the mollusc, Crepidula fornicata. Genesis. 53 (2), 237-244 (2015).
- Kandel, E. R. The molecular biology of memory storage: a dialog between genes and synapses. Bio. Rep. 24, 475-522 (2004).
- Jackson, D. J., Ellemor, N., Degnan, B. M. Correlating gene expression with larval competence, and the effect of age and parentage on metamorphosis in the tropical abalone Haliotis asinina. Mar. Biol. 147, 681-697 (2005).
- Carter, C. J., Farrar, N., Carlone, R. L., Spencer, G. E. Developmental expression of a molluscan RXR and evidence for its novel, nongenomic role in growth cone guidance. Dev. Biol. 343 (1-2), 124-137 (2010).
- Rittschof, D., McClellan-Green, P. Molluscs as multidisciplinary models in environment toxicology. Mar. Pollut. Bull. 50 (4), 369-373 (2005).
- Liu, M. M., Davey, J. W., Jackson, D. J., Blaxter, M. L., Davison, A. A conserved set of maternal genes? Insights from a molluscan transcriptome. Int. J. Dev. Biol. 58 (6-8), 501-511 (2014).
- Hohagen, J., Herlitze, I., Jackson, D. J. An optimised whole mount in situ. hybridisation protocol for the mollusc Lymnaea stagnalis. BMC Dev. Biol. 15 (1), 19 (2015).
- Raven, C. P. The development of the egg of Limnaea stagnalis. L. from oviposition till first cleavage. Arch. Neth. J. Zool. 1 (4), 91-121 (1946).
- Raven, C. P. The development of the egg of Limnaea Stagnalis. L. from the first cleavage till the troghophore stage, with special reference to its’ chemical embryology. Arch. Neth. J. Zool. 1 (4), 353-434 (1946).
- Raven, C. P. Morphogenesis in Limnaea stagnalis. and its disturbance by lithium. J. Exp. Zool. 121 (1), 1-77 (1952).
- Raven, C. P. The nature and origin of the cortical morphogenetic field in Limnaea. Dev. Biol. 7, 130-143 (1963).
- Morrill, J. B., Blair, C. A., Larsen, W. J. Regulative development in the pulmonate gastropod, Lymnaea palustris., as determined by blastomere deletion experiments. J Exp Zool. 183 (1), (1973).
- Van Den Biggelaar, J. A. M. Timing of the phases of the cell cycle during the period of asynchronous division up to the 49-cell stage in Lymnaea. J. Emb. Exp. Morph. 26 (3), 367-391 (1971).
- Verdonk, N. H. Gene expression in early development of Lymnaea stagnalis. Dev. Biol. 35 (1), 29 (1973).
- Gall, J. G., Pardue, M. L. Formation and Detection of Rna-Dna Hybrid Molecules in Cytological Preparations. Proceedings Of The National Academy Of Sciences Of The United States Of America. 63 (2), 378-383 (1969).
- Iijima, M., Takeuchi, T., Sarashina, I., Endo, K. Expression patterns of engrailed and dpp in the gastropod Lymnaea stagnalis. Dev Genes Evol. 218 (5), 237-251 (2008).
- Shimizu, K., Sarashina, I., Kagi, H., Endo, K. Possible functions of Dpp in gastropod shell formation and shell coiling. Dev Genes Evol. 221 (2), 59-68 (2011).
- Koop, D., Richards, G. S., Wanninger, A., Gunter, H. M., Degnan, B. M. D. The role of MAPK signaling in patterning and establishing axial symmetry in the gastropod Haliotis asinina. Dev. Biol. 311 (1), 200-212 (2007).
- Lartillot, N., Lespinet, O., Vervoort, M., Adoutte, A. Expression pattern of Brachyury in the mollusc Patella vulgata suggests a conserved role in the establishment of the AP axis in Bilateria. Development. 129 (6), 1411-1421 (2002).
- Jackson, D. J., Wörheide, G., Degnan, B. M. Dynamic expression of ancient and novel molluscan shell genes during ecological transitions. BMC Evol. Biol. 7 (1), 160 (2007).
- Jackson, D. J., Meyer, N. P., Seaver, E., Pang, K., McDougall, C., et al. Developmental expression of COE. across the Metazoa supports a conserved role in neuronal cell-type specification and mesodermal development. Dev Genes Evol. 220, 221-234 (2010).
- Perry, K. J., Lyons, D. C., Truchado-Garcia, M., Fischer, A. H. L., Helfrich, L. W., et al. Deployment of regulatory genes during gastrulation and germ layer specification in a model spiralian mollusc. Dev. Dyn. , (2015).
- Iijima, M., Takeuchi, T., Sarashina, I., Endo, K. Expression patterns of engrailed and dpp in the gastropod Lymnaea stagnalis. Dev Genes Evol. 218 (5), 237-251 (2008).
- Shimizu, K., Iijima, M., Setiamarga, D. H. E., Sarashina, I., Kudoh, T., et al. Left-right asymmetric expression of dpp in the mantle of gastropods correlates with asymmetric shell coiling. EvoDevo. 4 (1), 15 (2013).
- Christodoulou, F., Raible, F., Tomer, R., Simakov, O., Trachana, K., et al. Ancient animal microRNAs and the evolution of tissue identity. Nature. 463, (2010).
- Koga, M., Kudoh, T., Hamada, Y., Watanabe, M., Kageura, H. A new triple staining method for double in situ hybridization in combination with cell lineage tracing in whole-mount Xenopus embryos. Dev Growth Differ. 49 (8), 635-645 (2007).
- Lauter, G., Söll, I., Hauptmann, G. Two-color fluorescent in situ hybridization in the embryonic zebrafish brain using differential detection systems. BMC Dev. Biol. 11 (1), 43 (2011).
- Davison, A., Frend, H. T., Moray, C., Wheatley, H., Searle, L. J., Eichhorn, M. P. Mating behaviour in Lymnaea stagnalis. pond snails is a maternally inherited, lateralized trait. Biol. Lett. 5 (1), 20-22 (2009).
- Kuroda, R., Endo, B., Abe, M., Shimizu, M. Chiral blastomere arrangement dictates zygotic left-right asymmetry pathway in snails. Nature. 462 (7274), 790-794 (2009).
- Shibazaki, Y., Shimizu, M., Kuroda, R. Body handedness is directed by genetically determined cytoskeletal dynamics in the early embryo. Curr. Biol. 14 (16), 1462-1467 (2004).
- Lu, T. Z., Feng, Z. P. A sodium leak current regulates pacemaker activity of adult central pattern generator neurons in Lymnaea stagnalis. PLoS One. 6 (4), e18745 (2011).
- Dawson, T. F., Boone, A. N., Senatore, A., Piticaru, J., Thiyagalingam, S., et al. Gene Splicing of an Invertebrate Beta Subunit (LCav-beta) in the N-Terminal and HOOK Domains and Its Regulation of LCav1 and LCav2 Calcium Channels. PLoS ONE. 9 (4), e92941 (2014).
- Smith, S. A., Wilson, N. G., Goetz, F. E., Feehery, C., Andrade, S. C. S., et al. Resolving the evolutionary relationships of molluscs with phylogenomic tools. Nature. 480 (7377), 364-367 (2011).
- Gregory, T. R., Nicol, J. A., Tamm, H., Kullman, B., Kullman, K., et al. Eukaryotic genome size databases. Nuc. Acids. Res. 35 (Database issue), D332-D338 (2007).
