The Swiss rolling technique in combination with immunohistochemical staining allows for comprehensive analysis of small or large intestinal tissue. The example of H&E staining of a large bowel of a C57BL/6 mouse (Figure 1) is an illustration of the feasibility and the effectiveness of this technique. As shown in Figure 1, the image is able to capture all portions of the colon: proximal, middle, and distal. Thus, it allows for comprehensive histological assessment. The Swiss rolling and the ability to capture the entire length of the small or large intestinal tissue is extremely helpful for heterogeneous gene expression and marker staining or a variable response of the intestinal tissue to the insult.
An example is the EGFP staining pattern in Lgr5-EGFP/CreERT2 mice. In this mouse model, expression of EGFP is driven by the Lgr5 promoter, which is active only in the actively proliferating intestinal crypts. Additionally, this mouse model is characterized by low (approximately 5 – 10%) penetrance of transgene expression. As shown in Figure 2, the EGFP expression pattern in the intestinal tissue is variable. Consequently, the capability to capture a large view of the tissue helps to identify the region of interest.
The technique described here is powerful especially with application of multifluorophore staining. Here, we show an example of trifluorophore staining of the intestinal tissue that was prepared using the Swiss-roll technique. The main aim of this study was to investigate the role of Klf5 in the maintenance of intestinal stem cells expressing Lgr5 marker. Therefore, we deleted Klf5 from the Lgr5-positive intestinal stem cells in Lgr5-EGFP/CreERT2 mice and collected intestinal tissues on day 14 after first tamoxifen injection. For immunohistological analysis, the tissues were prepared according to the protocol presented in this manuscript, and staining for EGFP (Lgr5 marker), Klf5, and Ki-67 was performed. In the control mice, denoted as Lgr5-EGFP/CreERT2, both EGFP-positive (marked with blue arrows) and EGFP-negative crypts (marked with white arrows) exhibited co-staining for Klf5 and Ki-67 at two examined time points, as shown in Figure 3D. In contrast, in Lgr5-EGFP/CreERT2/Klf5fl/fl small intestinal tissue, Klf5/Ki-67 co-staining was missing from the EGFP-positive CBC stem cells (marked by blue arrows) but present in the EGFP-negative crypts adjoining the green crypts (marked by white arrows), as shown in Figure 3H. This is an excellent example that the tissue preparation techniques presented here does not negatively influence staining quality.
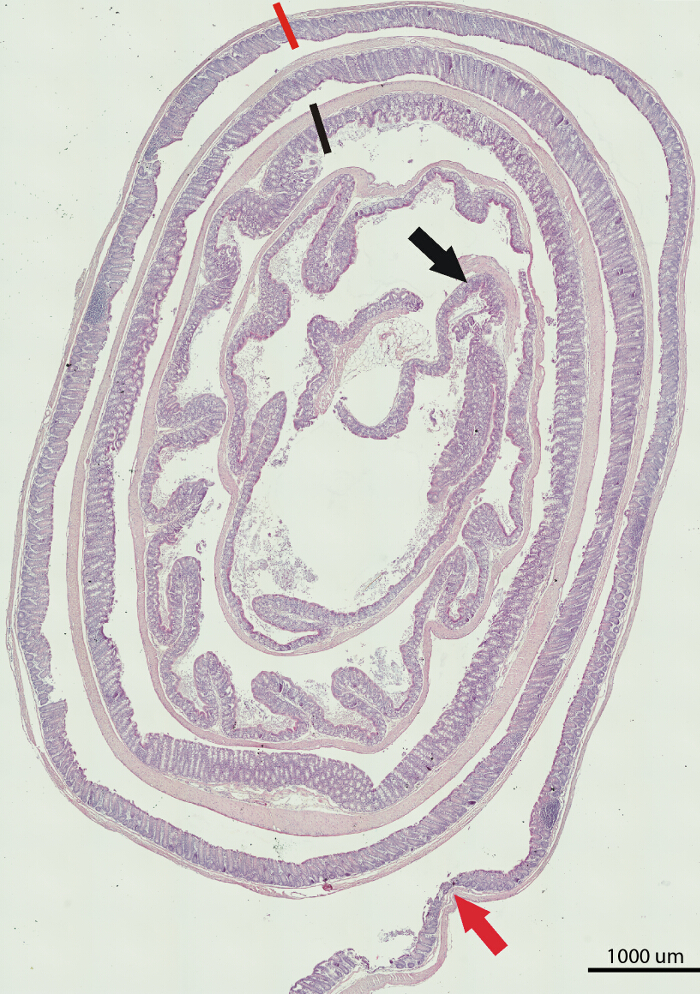
Figure 1. H&E Staining of a Large Bowel from a C57BL/6 Mouse. Shown is the composite image of the whole length of the large bowel. The portion between the black arrow and black line marks the proximal part, between the black and orange lines marks the middle, and between the orange line and orange arrow marks the distal part of the large bowel. Scale bar = 1,000 μm. Please click here to view a larger version of this figure.
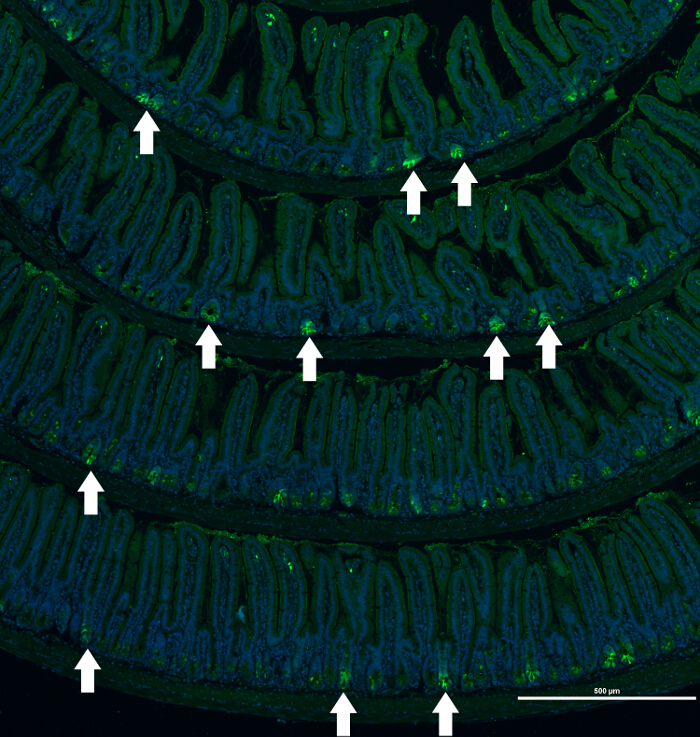
Figure 2. Immunofluorescence Staining of a Small Bowel of an Lgr5-EGFP/CreERT2 Mouse. Composite image of EGFP (labeling Lgr5-positive epithelial cells) staining of small intestine sections of an Lgr5-EGFP/CreERT2 mouse. Example of heterogeneous immunofluorescence staining of protein marker (EGFP) that labels Lgr5-postive epithelial cells in the crypts of the Lgr5-EGFP/CreERT2 mouse. Nuclei are visualized with Hoechst. White arrows mark crypts positive for EGFP expression. Scale bar = 500 μm. Please click here to view a larger version of this figure.
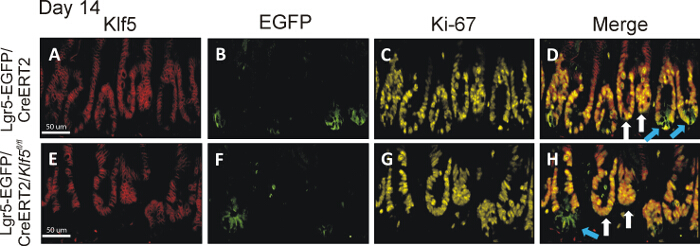
Figure 3. Klf5 Deletion in Lgr5-EGFP/CreERT2/Klf5fl/fl Mice Persists Long Term in Lgr5-EGFP-positive Crypts. The top and bottom set of images are representative small intestinal tissue from the Lgr5-EGFP/CreERT2 control and Lgr5-EGFP/CreERT2/Klf5fl/fl mice on day 14 day after first tamoxifen injection, respectively. Panels A-D are representative of staining from tissue collected from Lgr5-EGFP/CreERT2 control mice, while panels E-H show staining representative of Lgr5-EGFP/CreERT2/Klf5fl/fl mice. Panels A and E display Klf5 immunofluorescent staining in red; panels B and F show EGFP staining in green; panels C and G show Ki-67 staining in yellow; and panels D and H show merged images of Klf5, EGFP and Ki-67 stains. Blue arrows point to green crypts; white arrows point to nongreen crypts in the merged images. Lgr5-EGFP/CreERT2/Klf5fl/fl mice showed long-term loss of Klf5 only in the EGFP-labeled CBC cells14. Scale bar = 50 μm. Please click here to view a larger version of this figure.
