The x-y-z coordinates of the semi-landmarks produced in Protocol 6 can be directly imported into any landmark-based geometric morphometrics analysis17. The computational pipeline above has been applied to study mouse bacula14, as well as whale pelvic and rib bones16. More details on the computational definition of semi-landmarks are presented here, in an attempt to help researchers visualize steps that might be modified to accommodate their particular object of interest. The baculum contains several unique features that were exploited to automate certain transformations. For example, after computationally cutting the bone into two halves along a proximal-distal axis, we identified the proximal half simply by comparing the total number of points (the proximal has more). As long as there exist unique features such as this, our methods should be adaptable to any object. In addition, it should be emphasized that we empirically determined certain thresholds, such as "10% proximal" that performed well in our baculum studies, but most certainly need to be reassessed for other objects.
Beginning in Protocol 5, the first computational step is to calculate the convex hull (the smallest set of points that contains all other points in a sample) to identify the two points that are furthest apart from each other. These two points (red spheres, Figure 1C) begin to define a new z-axis (red line, Figure 1C) that runs proximal-distal through the bone. In the case of the baculum, the half of the point cloud that contains more points is defined as the proximal end.
Second, the entire point cloud is transformed so that the proximal point takes on the x,y,z coordinates of 0,0,0 and the distal point takes on the x,y,z coordinates of 0,0,+z, where +z is some positive value dependent on the size of the bone. At the end of this step, a z-axis passes through the length of the bone. For procedures below, the length from the minimum to maximum z-coordinates will be referred to as Zlength.
Third, to correct for variance associated with the exact placement of the proximal and distal points identified above, the 10% most proximal and 10% most distal points are sampled separately (Figure 1D), their respective centroids identified (red spheres, Figure 1E) and the point cloud transformed such that the proximal centroid is 0,0,0, and the distal centroid is 0,0,+z, with a new z-axis that passes through the center of the specimen (red line, Figure 1E).
Fourth, the point cloud is rotated around the z-axis by first taking a slice of points in the proximal 15 to 15.25% Zlength of the structure (blue points, Figure 1E). This slice of points is flattened in the z dimension (i.e. z-coordinates are simply ignored), the convex hull taken, and the minimum bounding rectangle (the smallest rectangle that contains all other points) calculated. Imagine a line connecting the midpoints of the two short sides of this minimum bounding rectangle. We rotate the point cloud until these two midpoints become -x,0,+z and +x,0,+z, respectively, thus this line becomes a new x-axis. After transformation, the distance between the maximal and minimal x values are referred to as Xlength. A new file is created from specimen.xyz to specimen.TRANSFORMED.xyz.
Fifth, points within 1% Xlength of the z-axis are sliced out (blue points, Figure 1F), and the single most proximal and single most distal point identified from this central slice and labeled DISTAL and PROXIMAL, respectively. These are the first two semi-landmarks identified.
Sixth, 50 evenly spaced slices of points are sampled along the z-axis (red points, Figure 1G). Each slice is a thickness of 1% Zlength. Each slice is then flattened in the z dimension, and divided equally by 7 vertical lines (red lines, Figure 1H). Points within 2% Xlength of each line are kept (red points, Figure 1H), then the points with the maximum and minimum y-coordinate are kept, projected onto each respective line, and labeled VENTRAL and DORSAL, respectively. In addition, labels contain the slice number and the line number, for example P15_VENTRAL4 is the ventral point sampled from the 4th vertical line of the 15th slice. Importantly, every point labeled, for example, P15_VENTRAL4, occurs once and only once across all specimens, preserving correspondence. In addition to the ventral and dorsal points of each of the 7 lines (14 semi-landmarks total), the points with the maximum and minimum x-value are sampled and labeled LEFT and RIGHT, respectively. The y and z coordinates of LEFT and RIGHT are smoothed using the lowess function in R. For the baculum, a total of 16 semi-landmarks are defined per slice (red spheres, Figure 1H); with 50 slices plus the PROXIMAL and DISTAL semi-landmarks defined above, 802 semi-landmarks are sampled per specimen (green spheres, Figure 1I). All other points from the original microCT scan are discarded.
It should be noted that although ventral/dorsal and proximal/distal polarity was determined mathematically, all specimen alignments were visually confirmed and manually adjusted as required. In our sample of 369 bacula, approximately 10 had to be manually adjusted.
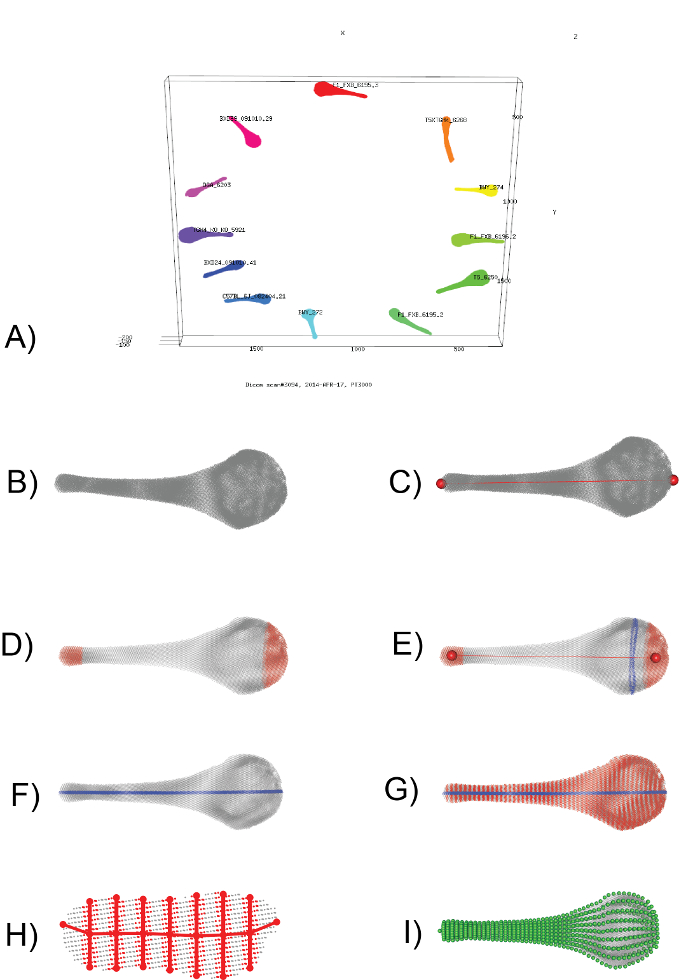
Figure 1: Visual Representation of the Computational Workflow (Protocol 4-6). (A) A screenshot from the 02_segment_dicoms.r script (Protocol 4), showing the assignment of distinct point clouds to individual specimens. (B) Enlarged view of one baculum, represented by a cloud of ~100K x-y-z points. (C) Identification of the two points furthest apart from each other (red spheres), used to define a new z-axis that runs proximal-distal through the bone (red line). (D) Sampling the proximal-most 10% and distal-most 10% of points (red points) provides a means to adjust for slight variance in the exact placement of the z-axis. (E) The centroids of the proximal-most 10% and distal-most 10% (red spheres) are used to define a new z-axis (red line). Then, a slice of points falling between 15.00-15.25% of this new z-axis (blue points) is taken to calculate the minimum bounding rectangle. The point cloud is rotated until the long side of the minimum bounding rectangle is parallel to a new x-axis. F) a slice of points running along the midline (blue points) is sampled and the proximal-most and distal-most point defined as a semi-landmark. G) 50 evenly spaced slices of points are sampled (red points), with H) showing one such slice. 16 points (red circles) are defined to capture the outline of each slice. I) When repeated across all slices, a total of 802 semi-landmarks (green spheres) define the structure and are used in all downstream morphometric applications. Please click here to view a larger version of this figure.
