To identify the location of two DC subsets, CD11bhi cDCs and CD103+ cDCs, PCLS from C57BL/6 mice were cut and stained with monoclonal antibodies (mAbs) specific to CD11c, CD88, CD103, and CD324 (E-cadherin). Antibodies to CD324 stain airway epithelial cells, and CD88 is displayed on macrophages and neutrophils, but not cDCs8. This allowed us to distinguish cDCs from CD11c+ macrophages, and to observe the spatial relationship of each cell type to the airways (Figure 1). We found that CD11bhi cDCs localize in the parenchyma, whereas CD103+ cDCs reside primarily around the airways and the subpleural area. Although CD103+ cDCs were readily detected by direct staining of cell surface CD103 in this experiment, the detection of CD11bhi cDCs relied on the absence of CD88 and CD103 staining. To directly identify CD11bhi cDCs, we first attempted to stain PCLS using an anti-CD11b mAb (clone M1/70), which is widely used in immunohistochemistry and flow cytometry. However, this mAb had a high background and low specificity in this application, regardless of which fluorochromes were conjugated to the mAb (Figure 2A, and data not shown). By contrast, antibodies to CD172a (SIRP1α) stained CD11bhi cDCs18, but not structural cells or CD103+ cDCs (Figure 2B, and data not shown). On their own, antibodies to CD172a could not discriminate between cDCs and monocytes or macrophages. However, by co-staining with antibodies against CD88 and CD172a, we were able to distinguish alveolar macrophages (CD172a–CD88+) from interstitial macrophages (CD172a+CD88+), and CD11bhi cDCs (CD172a+CD88–), and show that the latter cells preferentially localize in the parenchyma, not the sub-epithelial area (Figure 2C – F), in agreement with our results obtained by indirect staining of these cells.
Study of intra-tissue migration of leukocytes requires visualization of structural cells, including those of the lymphatics, through which DCs traffic from peripheral tissues to regional LNs. In the skin and LNs, lymphatic vessels are often identified using mAbs to LYVE-1 or Podoplanin. However, we found that in PCLS, mAbs to LYVE-1 and Podoplanin stain a variety of other cell types, including vascular endothelium and alveolar epithelium (data not shown). These mAbs are therefore of limited practical use for specifically identifying lymphatic vessels in PCLS. However, mAbs to CD90.2 (Thy1.2), a well-known T cell marker, labeled lymphatic structures, as shown previously in other tissues19,20 (Figures 3A, B), as did TdTomato fluorescent protein encoded by a transgene under transcriptional control of the Prox1 promoter21 (Figure 3A, C). PROX1 is a master transcription factor that is necessary for lymphangiogenesis22. Prox1TdTomato transgenic mice exhibit brightly fluorescent lymphatic vessels in the lungs and other tissues such as the liver, lens, dentate gyrus and neuroendocrine cells of the adrenal medulla21. Indeed, these two labeling methods gave overlapping staining patterns (Figure 3D). Using these labeling approaches, we were able to determine that CD103+ cDCs were present in several anatomical areas, including CD324-expressing airway epithelium, the lymphatics, and the subpleural area (Figure 3A, E).
In addition to static imaging, cell movement and cell to cell interactions can be recorded using live cell imaging of PCLS (Movie 1). PCLS were freshly prepared from mice that had received injections of GFP-expressing OT-II T cells and instillation of OVA and LPS, stained with mAbs against CD103 (red) and the airway epithelium (CD324, blue), and were imaged to track the movement of CD103+ cDCs and T cells 16 h after OVA/LPS instillation. Over the course of the video (4 h), the dynamic movement of CD103+ DCs (red) and the adoptively transferred OVA-specific T cells (green) and their interaction are clearly visible (Movie 1).
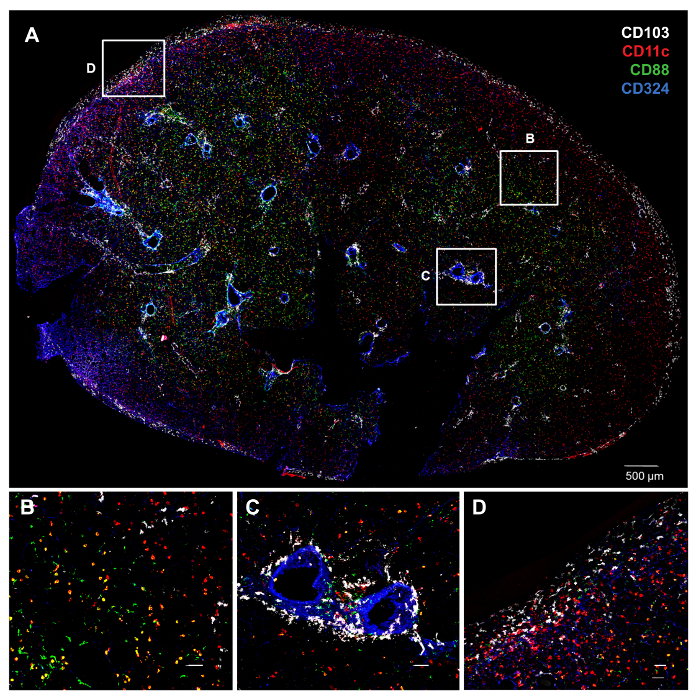
Figure 1. Distinct Anatomic Locations of cDC Subsets as Revealed by PCLS Staining and Confocal Microscopy. A) Whole PCLS from an untreated C57BL/6 mouse lung stained with various antibodies. The color of each molecule is indicated by its font color (top right). Individual and combinatorial staining gives the following colors for cell types of interest: CD103+ cDCs (CD103+CD11c+CD88–; white), CD11bhi cDCs (CD11c+CD103–CD88–; red), macrophages (CD11c+CD88+; yellow), neutrophils (CD11c–CD88+; green) and airway epithelial cells (CD324+; blue). B–D) Higher magnification of the insets in A. B) CD11bhi cDCs localize in the parenchyma. C) CD103+ cDCs around the airways. D) CD103+ cDCs underneath the visceral pleura. White bars denote 50 µm. Please click here to view a larger version of this figure.
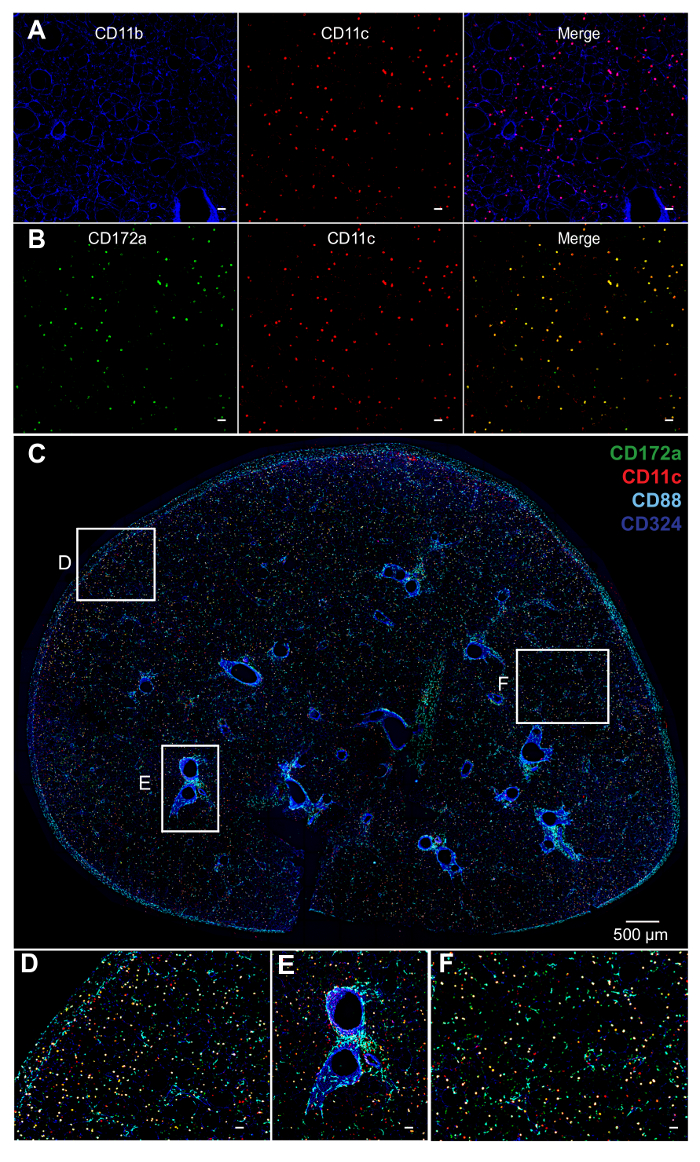
Figure 2. Staining of DCs and Macrophages. A) Staining of PCLS with antibodies to CD11b (blue) gives a high background in PCLS. B) Antibodies to CD172a (green) have a relatively low background in PCLS and greater specificity for CD11b+ cells. C) Whole PCLS of C57BL/6 mice stained with mAbs whose color is indicated by font color (top right). CD11c (red), CD88 (cyan), CD172a (green) and CD324 (blue). CD11bhi cDCs (CD11c+CD88–CD172a+; yellow), alveolar macrophages (CD11c+CD88+CD172a–; pink), interstitial macrophages (CD11c+CD88+CD172a+; white), neutrophils (CD11c–CD88+; cyan) and airways (CD324+; blue). D – F) Higher magnification of the insets depicted in C. D) Sub-pleural area. E) Sub-epithelial area. F) Interstitium. Interstitial macrophages localize in the parenchyma, and alveolar macrophages are in alveoli. Unlabeled white bars denote 50 µm. Please click here to view a larger version of this figure.
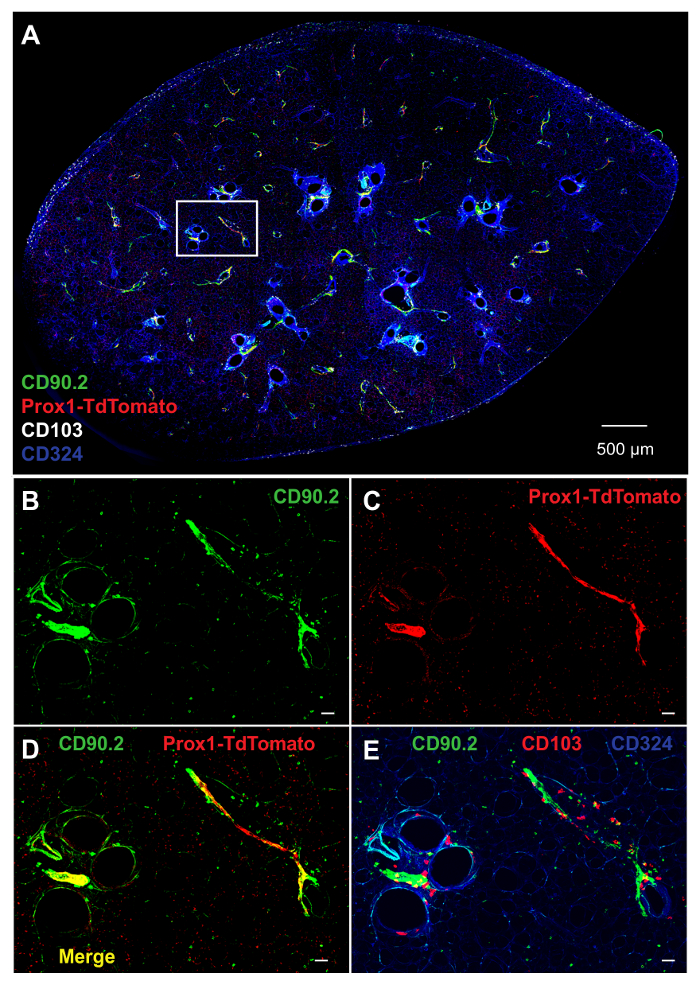
Figure 3. Labeling of Structural Cells in PCLS. A) Whole PCLS prepared from Prox1tdTomato mouse, and stained with mAbs to CD90.2 (green), CD103 (white) and CD324 (blue). Prox1-expressing cells (red) are genetically labeled with TdTomato under the regulation of Prox1 promoter (Prox1-TdTomato). B–E) Higher magnification of the inset in A. B) CD90.2 (green), C) Prox1-TdTomato (red), and D) combination of CD90.2 and Prox1-TdTomato. E) Combination of CD90.2 (green), CD103 (red) and CD324 (blue). Unlabeled white bars denote 50 µm. Please click here to view a larger version of this figure.
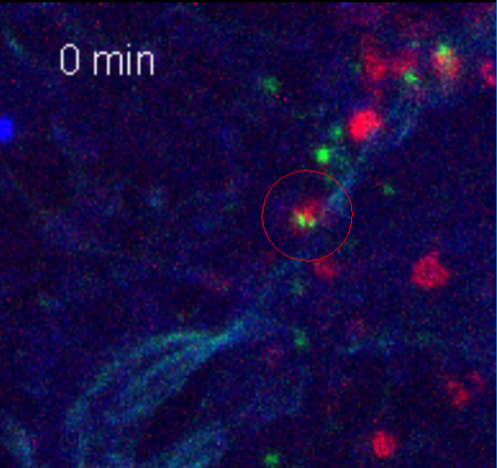
Movie 1. T Cells Interact with CD103+ cDCs in the Lung. Live cell imaging of CD103+ cDCs (red) interacting with T cells (green). CD4+ T cells isolated from OVA-specific OT-II Nur77gfp mice were stimulated with splenic DCs and OVA in vitro, and adoptively transferred into Rag -/- mouse. 2 h later, the recipient mouse was treated with OVA/LPS instillation to the airway. 16 h after OVA/LPS treatment, PCLS were made, stained and imaged for 4 h at 37 °C. Time recording image (min) is shown. Please click here to view this video. (Right-click to download.)
