Tumor transplantation and engraftment
A schematized outline of the procedure is represented in Figure 1. Fluorescently labeled tumor cells are extracted from the primary organ site, and a single-cell suspension is generated for transplantation into the fourth ventricle of 2-dpf mitfa w2 zebrafish embryos44. Figure 2 shows the representative results for this method using a zebrafish model of central nervous system primitive neural ectodermal tumor (CNS-PNET), in which the sox10 promoter drives expression of wildtype NRAS or activated NRAS fused to mCherry in p53-deficient oligoneural precursor cells17. The mitfa w2 zebrafish44 mutant, which lacks melanophores, were used to more readily visualize the tumor cells after transplantation and/or into adulthood. Other pigment mutants could also be used to provide additional imaging clarity, such as Casper50. As early as 24 h post-transplantation, tumor cells can be seen invading into surrounding brain tissue (Figure 2A, 3 dpf, arrows). Tumor grafts continue to grow in the ventricle and surrounding brain tissue over the next week (Figure 2A, 8 dpf). At this time, fluorescence can also be observed in the region of the kidneys (Figure 2A, asterisks). This may be due to the kidneys filtering the cellular debris from the transplantation, which is consistent with prior studies that use fluorescent dyes as an assay to evaluate kidney function51. Tumor cells continue to proliferate and invade, so by 28 dpf, a tumor mass can be seen throughout the zebrafish brain (Figure 2A, 28 dpf).
This technique allows for the rapid transplantation of tumor cells into hundreds of embryos with reproducible consistency, as the injection site can be targeted accurately with minimal damage to the host. A representative group of embryos transplanted with mCherry-labeled brain tumor cells is shown in the top panel of Figure 2B. Tumor engraftment is typically achieved in 80-90% of embryos, and tumor transplants persist into adulthood; three representative examples are shown in the bottom panels of Figure 2B. This allows for the tumor transplants to be harvested and frozen or re-transplanted over several generations.
Tumor invasion and drug responses
Figure 3A shows a co-transplantation scheme in which two different tumors labeled with either GFP (Tumor A) or mCherry (Tumor B) are harvested by whole-tumor dissociation (note that FACS is not used here, but FACS-sorted tumor cells could be used if needed). Tumor A and B can be from different locations, induced with different oncogenes, or induced in different genetic backgrounds. Each tumor is processed into a single-cell suspension, and tumor cells are then mixed together at a 1:1 ratio and transplanted into the 2-dpf embryo. Tumor properties, such as engraftment, growth, invasion, and dissemination of Tumor A versus Tumor B can then be observed in the same host that allows internal controls for the technique and genetic background.
In Figure 3B, an NRAS-driven, mCherry-labeled zebrafish CNS-PNET from the optic tectum and an GFP-labeled CNS-PNET from the cerebellum were harvested and processed into single-cell suspensions. Cells were counted, and an equal number of cells from each tumor were mixed together and transplanted into 2-dpf embryos. Four days after transplantation, the embryos were imaged with confocal microscopy. Shown is a representative embryo demonstrating that the tectum tumor cells (red) migrate more extensively into the host brain (arrows) than the cerebellar tumor.
Embryos transplanted with fluorescently labeled tumor cells can be used in drug treatment regimens to identify compounds that inhibit tumor growth. Figure 3C is a typical timeline for treating and imaging tumor transplants. 24 h post-transplantation, embryos are accessed for consistent engraftment size and split into treatment groups. In the provided example, embryos transplanted with NRAS-driven, mCherry-labeled zebrafish CNS-PNETs are treated with 50 µM dimethyl sulfoxide (DMSO) or MEK inhibitor (AZD6244). Embryos are treated for 5 days and imaged at 8 dpf, as outlined in Figure 3C. Figure 3D (left panels) shows representative animals from the two treatment groups. MEK inhibitor-treated embryos have a significantly lower amount of fluorescence than those treated with DMSO, as described17. After the animals are imaged, they are transferred to tanks without drugs and monitored for tumor growth for two months. In the provided example, the MEK inhibitor treatment results in a durable response in the adult fish (Figure 3D, right panels). For further details on the drug treatment, imaging, and quantification, see reference17.
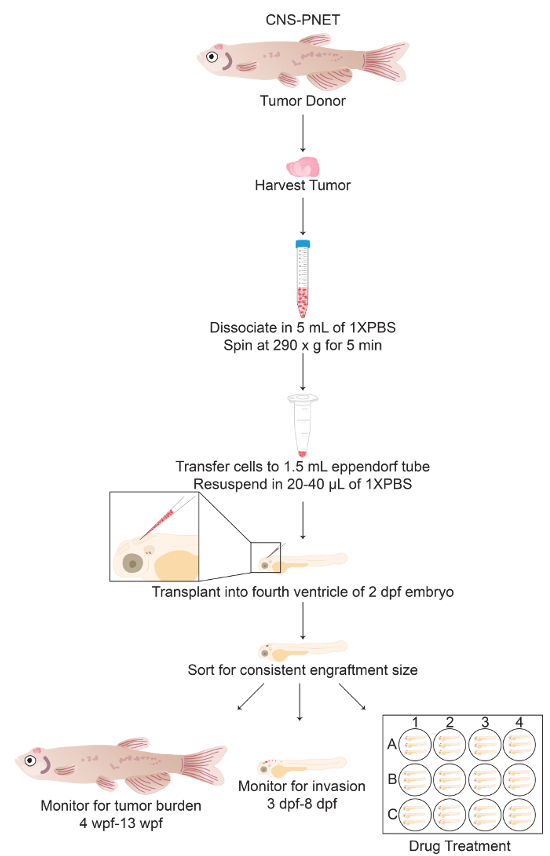
Figure 1: Protocol Schematic of Tumor Cell Transplantation into the Fourth Ventricle of 2-dpf Zebrafish Embryos. General schematic of sections 3 through 4 of the protocol including alternative endpoint analyses (sections 4.9-6). Please click here to view a larger version of this figure.
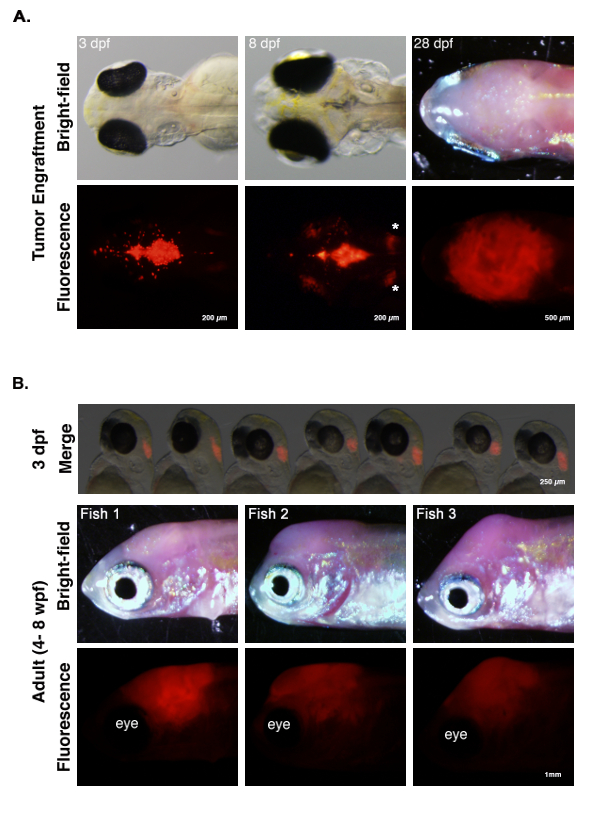
Figure 2: Tumor Engraftment and Progression. (A) Zebrafish primary CNS-PNET17 tumor cells labeled with mCherry were transplanted into the fourth ventricle of 2-dpf embryos. At 3 dpf, tumor cells are seen invading into surrounding brain tissue (arrows). At 8 dpf, tumor cells are growing in the ventricle; red tumor cells are also present in the surrounding brain tissue and kidneys (asterisks). At 28 dpf, tumors cells have invaded and grow throughout the host brain. (B) Top panel. Seven 3-dpf embryos transplanted with mCherry-labeled zebrafish brain tumor cells. Tumor cells can be transplanted consistently into hundreds of embryos, with high engraftment rates (i.e., typically 85/100 embryos). Middle and bottom panels. Three representative adult fish with transplanted tumors 4-8 weeks post-fertilization (wpf). Tumors can be visualized by fluorescence microscopy. Please click here to view a larger version of this figure.
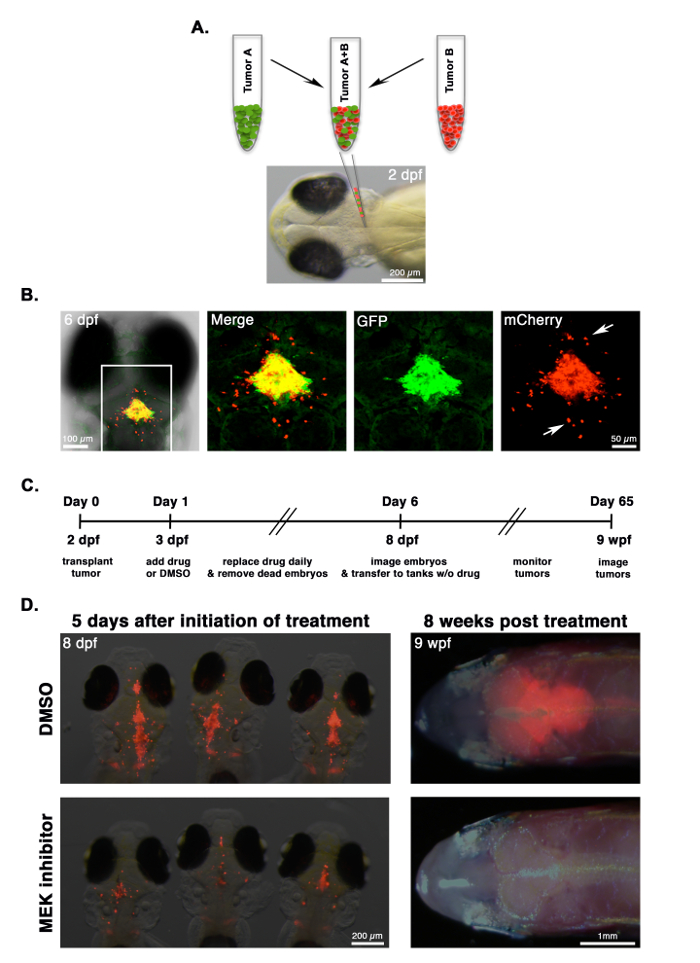
Figure 3: Comparing Tumor Invasion and Drug Response. (A) CNS-PNETs17 from the optic tectum or cerebellum are labeled with mCherry or GFP, respectively, processed into a single-cell suspension, mixed together at a 1:1 ratio, and co-transplanted into 2-dpf embryos. (B) A 6-dpf embryo co-transplanted with a mixed suspension of GFP-labeled cerebellar tumor cells and mCherry-labeled tumor cells from the optic tectum. Differences in the invasive properties of each tumor in the same recipient can be observed using fluorescence microscopy. In this example, mCherry-labeled tumor cells migrate more extensively than those labeled with GFP (arrows). (C) Timeline for drug treatment on embryos transplanted with tumor cells. (D) Images of representative animals at the end of the treatment regimen (8 dpf) and at 8 weeks post-treatment (8 wpf), with either a DMSO control or a 50 µM MEK inhibitor. Tumor burden can be measured by quantifying fluorescence, as described17. In this experiment, the growth of mCherry-labeled tumor cells is reduced in MEK inhibitor-treated embryos and translates into a durable response in the adult. Please click here to view a larger version of this figure.
