18F-FDG PET imaging was performed on KL mice and demonstrated that the tumors in these mice were highly glycolytic as shown by an elevated 18F-FDG consumption (Figure 1A), agreeing with previously published studies26,29. A resection of whole lungs revealed the presence of several tumors (Figure 1B). Mouse lungs can be divided up into 5 separate lobes represented in Figures 1C and 1D. Lobes 1 – 5 were labeled on sectioned lungs that were stained with H&E or glucose transporter 1 (Glut1) (Figure 1D). Glut1 is a primary transporter of both glucose and 18F-FDG and its expression and localization to the plasma membrane of tumor cells directly correlate with 18F-FDG SUV29. A higher resolution analysis of Glut1 staining (40X) in 18F-FDG-avid lung tumors shows an elevated expression and localization of the transporter at the plasma membrane (Figure 1D).
Due to the limited resolution of PET imaging, both PET/CT and tissue autoradiography were performed. The higher resolution of autoradiography might identify smaller tumors and/or heterogeneity of the tumor 18F-FDG distribution. After the tumor induction, 18F-FDG PET/CT imaging was performed on KL mice (Figure 2A) followed by autoradiography on lungs isolated from these mice (Figures 2B and 2C). As seen in Figures 2B and 2C, the autoradiography identified two additional smaller tumors that were positive for 18F-FDG yet were not readily visible by PET. Following autoradiography, the slides with tissue could also be used for the immunohistochemical (IHC) staining of biomarker(s). H&E staining for the tumors confirmed the presence of tumors in the left lobe (Figure 2D).
Next, 18F-FDG PET imaging was performed on MLN0128-treated KrasG12D;Lkb1-/- mice in order to utilize 18F-FDG as a functional biomarker of glucose metabolism in lung tumors (Figure 3). We identified that a treatment with MLN0128 robustly inhibited the mTORC1 signaling and glycolysis as shown by a reduced 18F-FDG consumption (Figures 3A and 3B). These results agree with preclinical studies assessing MLN0128 in KL mice as previously published by our laboratory17,27. Lastly, IHC staining was performed on the tumors (Figure 3C). Tumors were stained either for H&E or with antibodies against phospho-S6, which is a conserved substrate of mTORC1 and is used to indicate an mTORC1 activation (P-S6) vs. inactivation (S6). Figure 3C shows a robust inhibition of P-S6 by MLN0128 in KL tumors compared to those treated with a vehicle, which agrees with previously published work17. In addition to KRAS, oncogenic drivers such as the epidermal growth factor receptor (EGFR) support the glycolytic metabolism in lung tumors as well. Therefore, we tested whether the inhibition of constitutively active mutant EGFR with erlotinib suppressed the 18F-FDG metabolism in mouse xenografts. Figures 3D and 3E show that the human lung tumor line HCC827, which harbors an EGFR del19 mutation, showed a significantly reduced 18F-FDG consumption following five days of erlotinib treatment.
Lastly, morphometric tissue analysis was performed on sectioned lungs and lung tumors to quantify the total tumor burden as well as to differentiate tumor pathologies that included tissue subtype, necrosis, blood vessels from normal lung tissue, and air space. The KL GEMMs developed a complex and pathologically heterogeneous disease that presented itself with lung tumors of varying histopathologies. These include adenocarcinomas (ADC) and squamous cell carcinomas (SCC)-this heterogeneity makes the treatment of this cancer a formidable challenge. Figure 4A shows a single H&E stained lung lobe with two large tumors present. The higher magnification images shown in Figure 4B identify a normal lung, vessels, and airspace, and tumor necrosis as well as adenocarcinoma, characterized by a well-defined papillary structure and a squamous cell carcinoma. Figure 4C represents the pseudo-coloring of the lung lobe and tumor using Inform morphometric software. Figure 4D shows the percentages of normal lung, vessels, and individual pathologies such as tumor necrosis and tumor subtypes that segmented well-differentiated adenocarcinoma from squamous cell carcinoma.
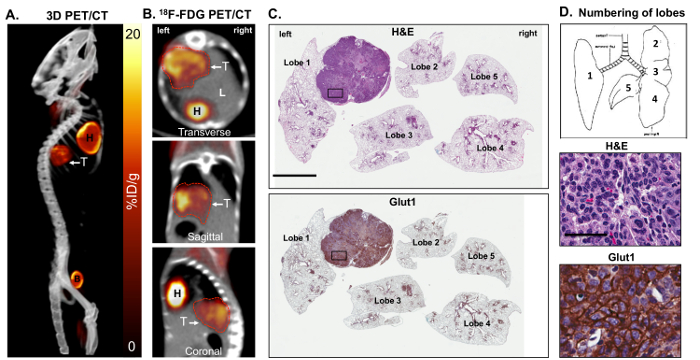
Figure 1: Metabolically active KrasG12D; Lkb1-/- (KL) mutant lung tumors are 18F-FDG positive and express high levels of glucose transporter 1 (Glut1). Panels A and B show a maximal intensity projection [also referred to as a 3-dimensional (3D) image] of the 18F-FDG-PET and CT analysis on some FDG-avid KL mice harboring squamous lung tumors. Shown are (A) a 3D reconstruction and (B) transverse, sagittal and coronal views of lung tumor(s) as (T). (C) This panel shows a whole lung histology of the KL mouse imaged in panels A and B, either stained for H&E (top panel) or with an antibody specific for Glut1 (bottom panel). The lung lobes are numbered. The scale bar = 2 mm. (D) This diagram represents the orientation and numbers of lobes in the mice (top panel) and higher resolution 40X images of the H&E- or Glut1-stained tumors from the slides shown in panel C for H&E (middle panel) or stained with an antibody specific for Glut1 (bottom panel). The scale bar = 25 µm. Please click here to view a larger version of this figure.
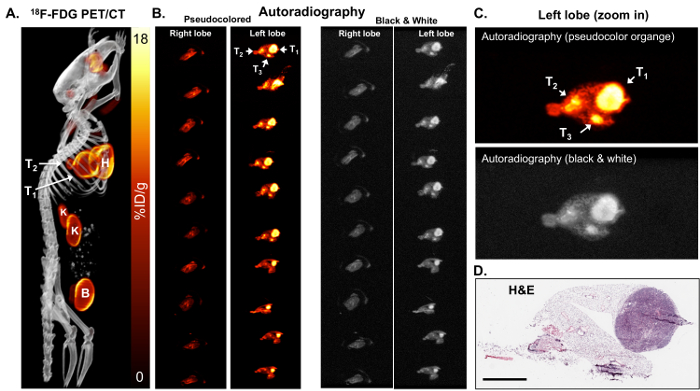
Figure 2: 18F-FDG autoradiography can identify small tumors that are metabolically active. (A) This 18F-FDG PET/CT image shows 18F-FDG-avid tumors in a KL mouse shown as a maximum-intensity projection image. T1 and T2 = tumors, H = the heart, B = the bladder, K = the kidneys. (B) This panel shows the ex vivo autoradiography on serial sections of the right and left lung lobes of the mouse. The lungs in the left and right panels are identical. The lungs in the left panels are pseudo colored orange. The lungs in the right panels are colored in black and white. The tumors (T1, T2, and T3) are indicated with arrows. (C) This is a magnified view of the autoradiography pseudocolored orange (top panel) and black and white (bottom panel). (D) This panel shows the H&E staining of the top slice of the left lobe shown in panel B. The scale bar = 200 µm Please click here to view a larger version of this figure.
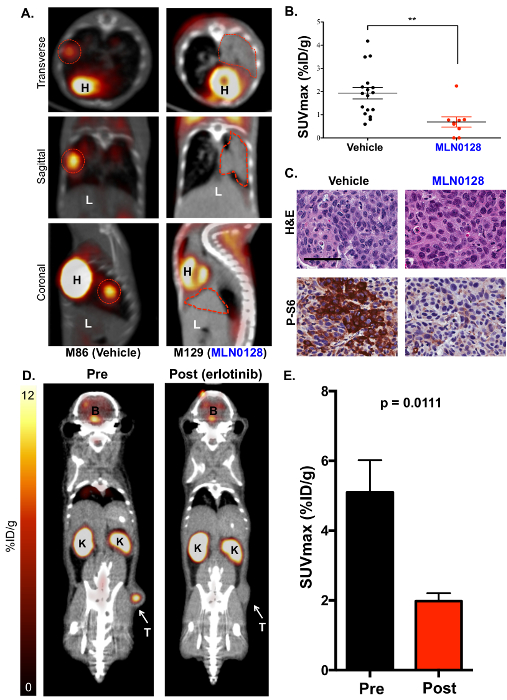
Figure 3: The mTOR inhibitor MLN0128 suppresses glucose consumption in lung tumors of KL mice as detected by 18F-FDG PET. (A) This panel shows representative 18F-FDG PET/CT images of KL mice treated with a vehicle (18F-FDG avid, left) or MLN0128 (18F-FDG non-avid, right). The transverse (top panel), coronal (middle panel), and sagittal (lower panel) views are shown. The tumors are outlined with red lines; H = the heart, L = the liver. (B) This panel shows a quantification of SUVmax (%ID/g) between the vehicle- and MLN0128-treated tumors. (C) This panel shows the H&E and P-S6 staining of whole lung sections from KL mice treated with vehicle or MLN0128. The scale bar = 25 µm. (D) This panel shows representative 18F-FDG-PET and CT images of HCC827 EGFR (del19) xenografts pre- and post-erlotinib treatment. The tumor (T) is indicated with an arrow, K = the kidney, B = the brain. (E) This panel shows a quantification of the SUVmax (%ID/g) for HCC827 xenografts before and after the erlotinib treatment. n = 10 tumors/group. Please click here to view a larger version of this figure.
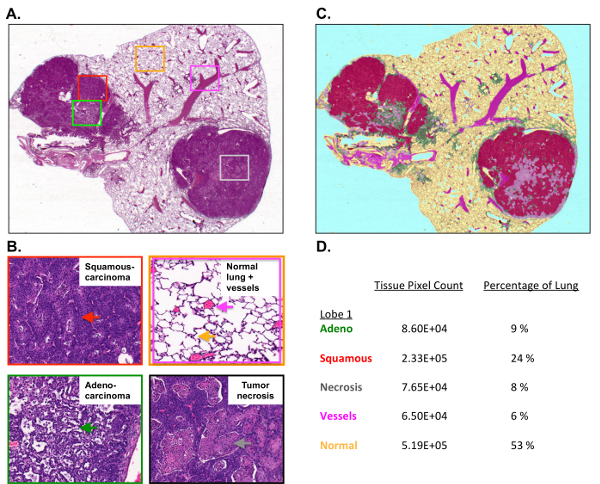
Figure 4: Tumor burden and tumor histology are quantified using morphometric software.
(A) This panel shows the H&E staining of a single mouse lung lobe with a tumor collected from a KL mouse. (B) These higher resolution images show the squamous cell carcinoma (top left), the normal lung, the vessels, and the air space (top right), and the well-differentiated papillary adenocarcinoma (bottom left) and necrosis (bottom right). (C) This panel shows the pseudocoloring of the H&E-stained lung lobe using morphometric software. (D) This panel shows the percentages for the individual lung lobe and tumor pathologies measured by Inform. Please click here to view a larger version of this figure.
