As illustrated in Figure 1, tip-growing plant cells encounter a series of physical barriers along their growth paths in vivo. The microfluidic in vitro cell culture platforms presented in this study enabled the examination the of tip-growing process in three types of plant cells (pollen tubes, root hairs, and moss protonemata) through 1 µm artificial gaps (Figure 3, Figure 4, Figure 5). For the pollen tube study, live-cell imaging was used to monitor morphological changes in the apical region of pollen tubes as well as the vegetative nucleus and sperm cells in response to encountering an extremely small space (Supplemental Movie 1 and 2).
Although most of the microgaps were successfully fabricated by employing the method described here, we noticed that a few of them were completely closed (Figure 6). Because the 1 µm wide channels created on the silicon mold are fragile, repeated use of this mold might damage the gaps, leading to gap blocking on the PDMS layer. Therefore, before performing the experiment, it is important to check that the PDMS microgaps are intact.
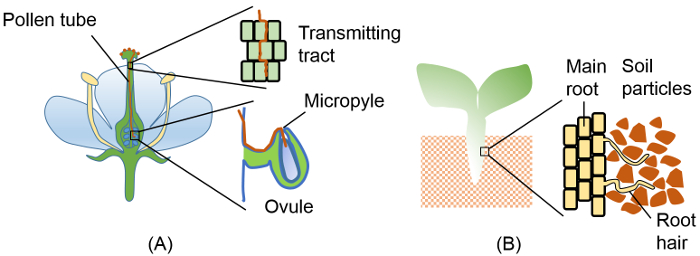
Figure 1: Penetration of tip-growing plant cells into physically constrained spaces. (A) Pollen tube elongating through several physical barriers on its way to an ovule. Barriers include transmitting tract tissue in the style and the integuments surrounding the micropyle. (B) Root hairs penetrating through dense soil. Insets in (A) and (B) show enlargements of the boxed regions. Please click here to view a larger version of this figure.
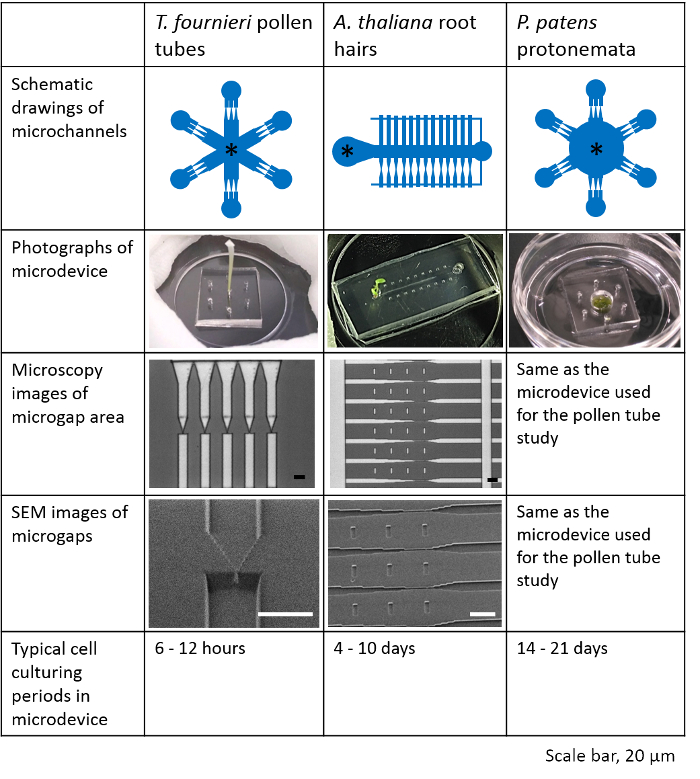
Figure 2: Characteristics of the microdevices used to study T. fournieri pollen tubes, A. thaliana root hairs, and P. patens protonemata. In the schematic drawings of microchannels, a sample is placed at the location indicated by an asterisk mark (*). Microchannel designs, photographs of each device, channel structures, and cell culturing periods are summarized. This figure has been modified with permission from Yanagisawa et al. 201719. Scale bar, 20 µm. Please click here to view a larger version of this figure.
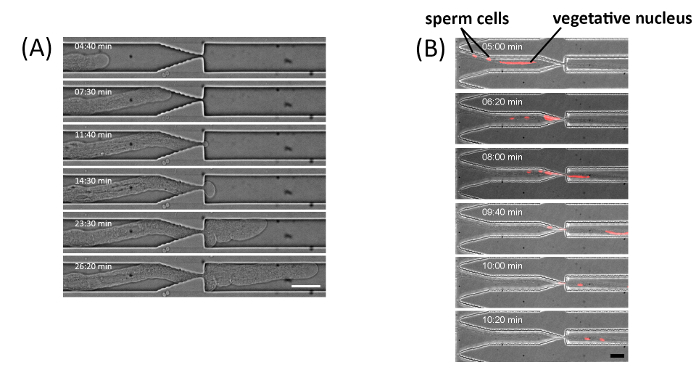
Figure 3: T. fournieri pollen tube elongation through a 1 µm gap. (A) Pollen tube passing through a 1 µm wide and 4 µm high gap. The time-lapse bright field images were captured using an inverted microscope equipped with a spinning-disk confocal system. Scale bar, 20 µm. (B) Time lapse images of a fluorescently labeled vegetative nucleus and sperm cells in a pollen tube of the RPS5Ap::H2B-tdTomato line crossing the microgap. Scale bar, 20 µm. Figure (A) and (B) have been modified with permission from Yanagisawa et al. 201719. Please click here to view a larger version of this figure.

Figure 4. A. thaliana root hair elongation through a 1 µm gap. (A) Root and root hair growth in the microdevice. The microgaps (1 µm in width and 4 µm in height) are located on the left side of the root growth channel. The microchannels on the right side do not contain microgaps and can be used as a control. Scale bar, 100 µm. (B) Bright field, (C) fluorescence, and (D) merged images of root hairs passing through the microgaps. The nuclei in root hairs are fluorescently labeled in the UBQ10pro::H2B-mClover line. The tip of each root hair is indicated by an arrow. Scale bar, 30 µm. These figures have been modified with permission from Yanagisawa et al. 201719. Please click here to view a larger version of this figure.
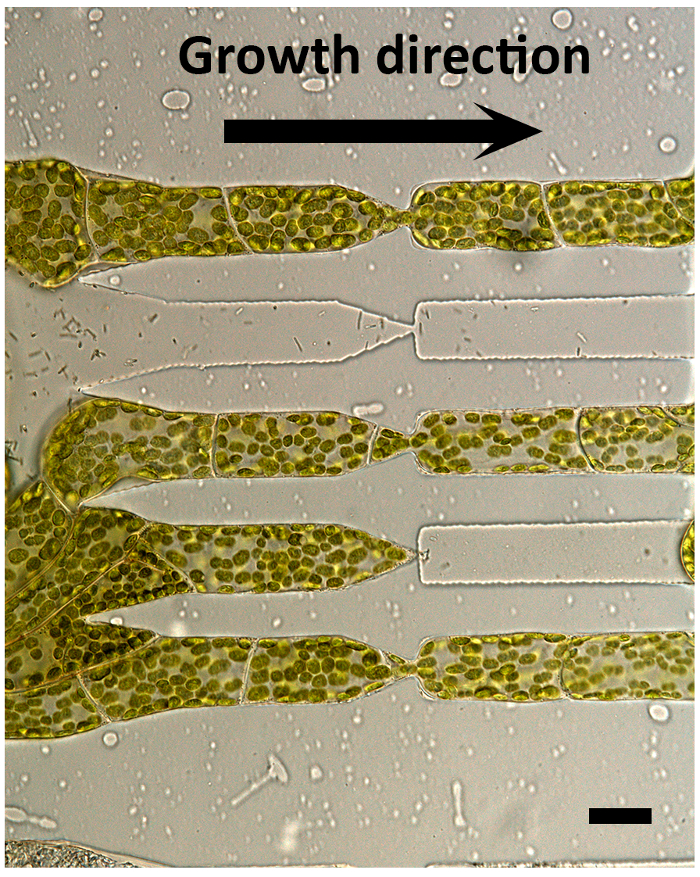
Figure 5. P. patens protonemata elongation through a 1 µm gap. Moss protonemata cells crossing through a 1 µm gap (4 µm in height), which was widened due to turgor pressure of the protonemata. Scale bar, 20 µm. The figure has been modified with permission from Yanagisawa et al. 201719. Please click here to view a larger version of this figure.

Figure 6. Closed microgaps. SEM image of the 1 µm gap region. The microgap on the right is completely closed. Please click here to view a larger version of this figure.
Supplementary Movie 1. Pollen tube growth through a 1 µm gap.Time-lapse bright field images were captured every 10 s using an inverted microscope equipped with a spinning disk confocal microscope. Scale bar, 20 µm. Please click here to download this file.
Supplementary Movie 2. Penetration of the vegetative nucleus and sperm cells through a 1 µm gap. Time-lapse bright field and fluorescence images were captured every 20 s using an inverted fluorescence microscope. Scale bar, 20 µm. Please click here to download this file.
