Impairment of the rec-8 Meiotic Cohesin Component Function Results in Diploid Gametes:
Imaging of the meiotic divisions of cohesin component mutant rec-8 sperm and oocytes revealed possible mechanisms for generating tetraploid animals (Figure 1)27,29,33. Mechanistically, the meiotic division defects of rec-8 mutant sperm and oocyte are different; however, both male and female diploid gametes are produced by the rec-8 mutant.
In wild type meiosis, homologous chromosomes become temporarily connected to one another by crossover recombination and enter the first meiotic division as a unit or bivalent (Figure 1A)34. During the first division, homologous chromosomes (homologs) segregate away from one another, whereas sister chromatids in each homolog remain together until the second division. Although the pattern of chromosome segregation is the same in female and male gametes, oocyte divisions are asymmetric whereas spermatocyte divisions are symmetric and undergo a specialized cytokinesis. In each division, the oocyte discards half of the division product into a small polar body. Thus, each oocyte precursor gives rise to a single haploid oocyte and two polar bodies (Figure 1B, C)35. Conversely, a single spermatocyte precursor gives rise to four functional spermatids by undergoing two symmetric divisions. The second division culminates with four spermatids budding off from a residual body. This cytokinesis is special in that the pattern and number of spermatids is determined by the centrosomes36.
In the precursor to the rec-8 mutant gametes, crossover recombination between homologous chromosomes does not take place and homologs are not connected as a bivalent at the beginning of the first meiotic division29,34. Sister chromatids of each homolog segregate away from one another in the first division instead of remaining together until the second division, as they do in wild type gametes. Interestingly, the rec-8 mutant phenotype during the second division is different in male and female gametes in that oocytes fail to undergo the cytokinesis whereas spermatocytes undergo a relatively normal cytokinesis (Figure 1B–F)27,28,29. Diploid oocytes arise because in the second division they fail both chromosome segregation and cytokinesis, hence they do not extrude the second polar body (Figure 1B, C). Spermatocytes in rec-8 germlines undergo spermatid budding or cytokinesis, but often segregate both sets of chromosomes to one of the spermatids to yield a diploid spermatid and a spermatid lacking chromatin (Figure 1D–F)27. Quantification of the proportion of rec-8 sperm lacking chromatin is shown in Figure 1F.
The formation of female and male diploid gametes in rec-8 mutants suggested that this mutant phenotype could be potentially used to generate full genome tetraploid strains.
Generation of Tetraploid C. elegans Strains:
The presence of a mutation in the rec-8 gene in all the generated tetraploid strains can be avoided by transiently knocking down rec-8 by RNAi29,37. This assumes that the reduction of rec-8 function by RNAi would generate diploid gametes that could potentially give rise to whole genome tetraploid animals, as rec-8 mutants do29,37. Essential to this protocol is that rec-8 mutants both give rise to diploid gametes and sire a reasonably large number of young, contrary to other meiotic mutants, which give rise to aneuploid gametes and are mostly sterile or embryonic/larval lethal.
Multiple tetraploid strains were generated by feeding C. elegans bacteria expressing rec-8 dsRNA using either one of two strategies (see Protocol, Figure 2, and Table 1)27. Tetraploids can be generated by self-fertilizing hermaphrodites for two to three generations in Petri dishes with bacteria expressing rec-8 dsRNA. By this strategy a hermaphrodite is placed in freshly made rec-8 RNAi plates and its progeny is transferred a few days later onto new plates with freshly induced rec-8 RNAi expressing bacteria (see Protocol). In addition, tetraploids can be generated through crossing the first generation of hermaphrodites fed bacteria expressing rec-8 dsRNA with untreated males of the same genotype in Petri dishes with bacteria expressing rec-8 dsRNA (Figure 2). In this case, the males in the cross are exposed to the rec-8 RNAi bacteria from the L4 stage onwards, during mating. Both schemes give rise to tetraploid animals27. Table 1 shows tetraploid strains obtained using the scheme presented here. Triploid and tetraploid C. elegans are larger in size than diploids, but triploid strains are unstable and tend to become diploid in one or two generations, whereas tetraploid strains are relatively stable22,23. Putative tetraploid strains were identified as larger than the original strain (Lon) hermaphrodites that sired only Lon progeny (Figure 3A). Lon animals are easily identified – wild type diploid animals are two thirds of the length of tetraploids and their bodies do not make an extra bend, which can be noticed during its forward sinusoidal movement (Figure 3A).
Tetraploid strains were confirmed by screening for the presence of 12 chromosome pairs in oocytes of the tetraploid hermaphrodites compared to six chromosome pairs in oocytes of diploid hermaphrodites (Figure 3B, C, and Movie 1).
Tetraploid Classes:
Two types of tetraploid hermaphrodites were identified: one sired males at similar frequencies as diploid hermaphrodites and the other sired males at much higher frequencies (Table 1). These two kinds of tetraploid hermaphrodites have been shown to differ in that the class producing frequencies similar to those of diploid males are tetraploid for all its chromosomes (4A, 4X), whereas the class producing high frequencies of males are hermaphrodites that are tetraploid for the autosomes but triploid for the sex chromosome (4, 3X). The later class of tetraploids are stable and produce 4A, 3X hermaphrodites and 4A, 2X males.
Tetraploid strains grow slower and produce reduced brood size compared to the diploids they were derived from, as seen for the strains generated with the previous method22,23. Madl and Herman22 suggested that the increased proportion of dead embryos in the tetraploid strains could be due to aneuploidy in the oocyte, however superficial inspection of tetraploid strains did not reveal sufficiently elevated numbers of aneuploid oocytes nor abnormal oocyte or spermatocyte divisions to account for the observed reduction in brood size (Movie 2, and Figure 3C, D).
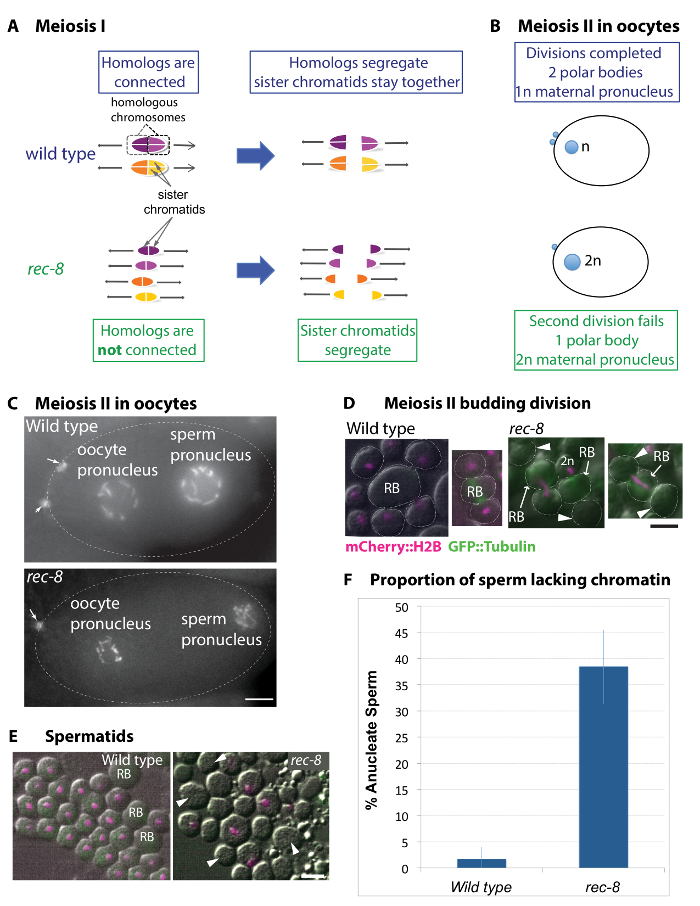
Figure 1: Gametogenesis in rec-8 mutant implies possible mechanisms for generating stable polyploid stains. (A) Diagram of the chromosome organization and segregation pattern in wild type and rec-8 mutant meiotic divisions. In wild type meiosis, homologous chromosomes separate in the first meiotic division. Sister chromatids in each homolog orient towards the same spindle pole and remain together until the second division. In rec-8 mutants, homologs do not form crossovers, and thus are not connected. In contrast to wild type, rec-8 sister chromatids orient away from one another and separate in the first meiotic division. (B) Diagram of wild type and mutant oocytes showing the female pronucleus and extruded polar bodies (male pronucleus is not depicted). In wild type oocytes, two asymmetric meiotic divisions result in the extrusion of two polar bodies. In rec-8 mutants however, the second polar body extrusion fails resulting in a diploid oocyte. (C) Images of wild type and rec-8 mutant oocytes expressing mCherry::histone H2B. Arrows indicate two polar bodies in the wild type oocyte and one in the rec-8 mutant. Scale bar is 5 µm. (D and E) Live Images of spermatids from wild type and rec-8 mutant animals expressing mCherry::histone H2B (magenta). Arrowheads indicate anucleate spermatids. Scale bar is 2 µm. (D) Spermatocytes undergoing the second (budding) division in the wild type and rec-8 mutant. The wild type spermatocytes undergo symmetric divisions resulting in four budding spermatids, each with a haploid chromosome complement. In rec-8 mutant spermatocytes, chromosome segregation is impaired in the second division. Most often a single mass of chromatin remains in the residual body (RB) or in one of the two sister spermatids in the second meiotic division. This gives rise to anucleate rec-8 mutant sperm (indicated by arrowheads) or diploid sperm. (E) Live images of post-budding spermatids in wild type and rec-8 mutants visualized using differential interference contrast (DIC) and fluorescence microscopy. Wild type spermatids all have similar chromatid masses. rec-8 mutant spermatids form anucleate sperm. (F) Quantification of wild type and rec-8 mutant anucleate sperm. rec-8 mutants produced 38.5% of anucleate sperm compared to less than 1.6% in wild type. Fisher's Exact test indicates that a rec-8 mutant has significantly higher incidence of anucleate sperm (p ≤0.0001) compared to the wild type. Error bars represent standard deviations. Please click here to view a larger version of this figure.
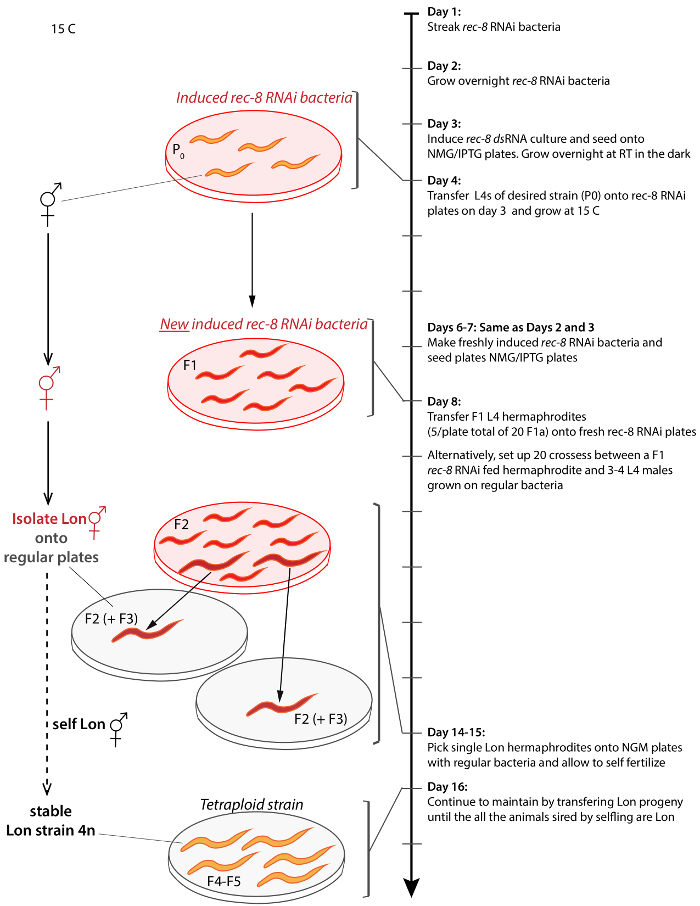
Figure 2: Scheme for generating and isolating tetraploid C. elegans from any strain. Arrow on the right represents the timeline of protocol starting from Day 1 and progressing to Day 17. Similar results can be achieved by crossing hermaphrodites to untreated males on Day 9. Brackets connect the depiction of a step to the timeline. Untreated animals are orange, treated animals are red, Lon animals are larger in size, rec-8 dsRNA expressing bacteria is depicted as transparent plates with transparent red background and regular OP50 bacteria is depicted on transparent grey background plates. Procedure is done at 15 °C unless otherwise stated. Please click here to view a larger version of this figure.
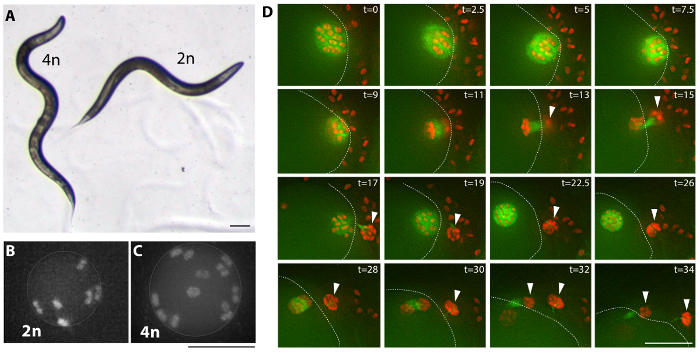
Figure 3: Tetraploid examples. (A) Bright field image of a tetraploid (MSC2) animal and the diploid strain it was derived from (AV740). Tetraploid C. elegans are overall larger and longer (Lon) than the diploid. This difference in body size results in an additional sinusoidal curve of the body of the moving tetraploid and can be used as a criterion to screen for tetraploid derivatives. Scale bar is 0.1 mm. (B, C) Fluorescence images of diploid (AV740) and tetraploid (MSC1) most mature unfertilized oocyte nulcei, prior to the meiotic divisions. Secondary screening is performed by observing the number of chromosome pairs in unfertilized oocytes of the established Lon strains, as shown in Movie 1 for an MSC1 strain expressing Mcherry::H2B and GFP::β-Tubulin in the germline. Scale bar is 5 µm. (D) Images from a time lapse (Movie 2) of tetraploid oocyte meiotic divisions depicting a generally normal meiosis. Arrow heads mark polar bodies and a dotted line marks the cortex of the oocyte inside the spermatheca and surrounded by sperm; t = time lapsed in minutes. Scale bar is 10 µm. Please click here to view a larger version of this figure.
| Genotype | Tetraploid derivative (sets of Autosomes : # of X-chromosomes)* |
Parental Strain (diploid) |
| meIs16 [pie-1p::mCherry::his-58 + unc-119(+)]; | MSC1 (4A:4X) | MSC0 |
| ruIs57 [pie-1p::GFP::b-tubulin + unc-119(+)] | MSC2 (4A:3X) | |
| MSC3 (4A:4X) | ||
| MSC5 (4A:3X) | ||
| MSC6 (4A:4X) | ||
| MSC8 (4A:4X) | ||
| unc-119(ed3) III; ddIs6; ddIs6[tbg-1::GFP + unc-119(+)]; ltIs37; ltIs37 [pAA64; pie-1p::mCherry::HIS-58 + unc-119(+)] IV | MSC14 (4A:3X) | TMR17 |
| MSC15 (4A:3X) | ||
| MSC16 (4A:4X) | ||
| spo-11(me44)/nT1 IV; +/nT1[ qIs51[myo-2::gfp Ppes-10::gfp, PF22B7.9::gfp]] V | AV800& | AV776 |
| meIs8 [pie-1p::gfp::cosa-1 + unc-119(+)] II; | AV809& | AV727 |
| ltIs37 [pie-1p::mCherry::his-58 + unc-119(+)] IV; | ||
| ltIs38 [pie-1p::gfp::ph(PLC1delta1) + unc-119(+)] | ||
| meIs8 [pie-1p::gfp::cosa-1 + unc-119(+)] II; mnT12 (X; IV) | AV826& | AV695 |
| mIn1[ dpy-10(e128) mIs14[myo-2::gfp pes-10::gfp]] / | AV810& | DR2078 |
| bli-2(e768) unc-4(e120) II | ||
| ruIs32 [pie-1::GFP::H2B + unc-119(+)] III | AV822& | AZ212 |
| AV823& | ||
| C. briggsae – mfIs42[Cel-sid-2 + Cel-myo-2::DsRed] | AV824& | JU1018 |
Table 1: Tetraploid strains generated. *Deduced by scoring the number of bivalents (connected homologous pairs) and univalents (individual homologs) in oocytes prior to the meiotic divisions in addition to the proportion of males sired.
Movie 1: Screening to confirm whether stable Lon strains are full or partial tetraploids. Movie starts with a diagram of a wild type hermaphrodite highlighting the region imaged (unfertilized oocytes) to identify tetraploids by counting chromosome pairs. Following the diagram, a series of Z-stack movies and projections show gonads of diploid and tetraploid animals fixed and DAPI stained or live images of strains expressing Mcherry::H2B histone. Screening is done by counting the number of connected homologous chromosome pairs in unfertilized oocytes. Counting of either MCherry or DAPI stained bodies is done in the unfertilized oocyte closest to the sperm storing spermatheca (the "-1 oocyte"). Chromosomes in this oocyte are most condensed and separated from one another, allowing for more accurate homolog pairs counts. More than 10 animals per strain were screened to ensure the -1 oocyte chromosome counts were accurate. Stack thickness is 0.2 µm. Please click here to view this video. (Right-click to download.)
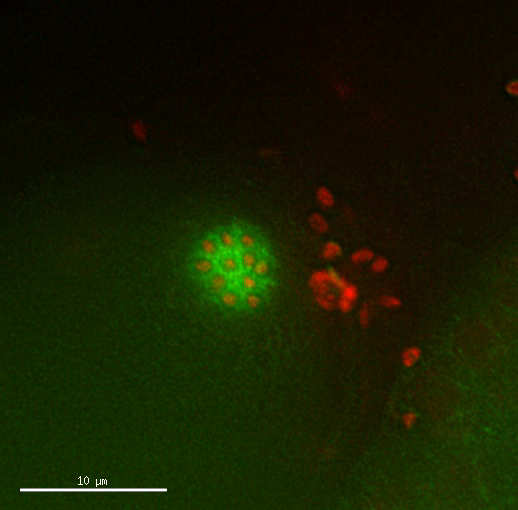
Movie 2: Tetraploid oocyte divisions appear normal. Time lapse of dividing oocyte of a tetraploid strain expressing Mcherry::H2B histone and GFP::β-Tubulin. Timing and pattern of the divisions appear normal. Images taken every 2.5 min. Please click here to view this video. (Right-click to download.)
