1. Media Preparation
- Buffered minimal media (BMG with glycerol, BMD with dextrose as carbon source) preparation (1 L)
- Autoclave 650 mL of distilled water in a 1 L screw cap glass bottle. After cooling down, complete the medium by adding 100 mL of autoclaved 10x YNB (134 g/L yeast nitrogen base w/o amino acids), 200 mL of potassium phosphate buffer (1 M, pH 6), 30-100 mL of 10% w/v glucose or glycerol (depending on the desired generated biomass amount during the batch), 2 mL of sterile 500x Biotin (10 mg/mL) solution and fill up to 1 L with sterile distilled water.
Note: All ingredients have to be autoclaved in separate flasks. Biotin is destroyed when autoclaved, and therefore has to be filter sterilized with 0.2 µm membrane filters.
- Autoclave 650 mL of distilled water in a 1 L screw cap glass bottle. After cooling down, complete the medium by adding 100 mL of autoclaved 10x YNB (134 g/L yeast nitrogen base w/o amino acids), 200 mL of potassium phosphate buffer (1 M, pH 6), 30-100 mL of 10% w/v glucose or glycerol (depending on the desired generated biomass amount during the batch), 2 mL of sterile 500x Biotin (10 mg/mL) solution and fill up to 1 L with sterile distilled water.
- Buffered rich media (BYPG) (1 L)
- Autoclave 700 mL of distilled water with 1% w/v yeast extract and 2% w/v peptone in a 1 L screw cap bottle. After cooling down, add 30-100 mL of the separately autoclaved 10% w/v glycerol, 200 mL of potassium phosphate buffer (1 M, pH 6) and fill up to 1 L with sterile distilled water.
2. Shake Flasks Preparation
- Add 5 mL of distilled water in the 250 mL baffled shake flasks, cover it with two layers of cotton cloth and fix it in place with rubber bands. Cover the cloth with a piece of aluminum foil.
- Autoclave the flasks at 121 °C for 20 min for full sterilization.
- Remove residual water from the flask and aliquot 50 mL of the previously prepared media to each flask.
3. Overnight Culture Inoculation
- Start an overnight culture (ONC) with a single fresh colony of the desired expression strain in 5 mL of YPD (1% w/v yeast extract, 2% w/v peptone, 2% w/v glucose) in a 50 mL centrifuge tube while leaving the lid slightly open to allow proper aeration.
- Let it grow over night at 28 °C, 80% humidity, in a shaker at 100-130 rpm (2.5 cm shaking diameter).
4. Measurement Setup and Main Culture Cultivation
- For setting up the online monitoring system, follow the manufacturer’s instructions for appropriate calibration of the devices.
- Place a computer, where the appropriate software for monitoring is installed in range for a Bluetooth connection to the measurement stations. Start the software to connect the devices via Bluetooth.
- Turn on the Bluetooth connection on the biomass measuring device by pressing the silver button on the front.
- In the software, click Devices then Find Devices.
Note: The devices should appear as a list in the window below the task line. If they do not, it is advisable to check if the batteries are charged and the computer is within reach for a Bluetooth connection. - Drag and drop the found devices to the squares on the right side of the screen Measurement Tray.
- Click Connect.
Note: When the connection was successful, a control screen appears. - To start a test run for the Bluetooth connection, click Measurement.
- Set up the experiment.
- Click Start Measurement, name the experiment and enable all parameters needed to monitor.
Note: Licenses are needed for each of the possible measured parameters. - Set Interval to 3 min.
Note: The interval determines how often measurement points are taken and every measurement point results in a value afterwards. Measuring every minute is very accurate, but lots of data are generated which makes it more laborious to evaluate. - Set Average Measurement Points to 11.
Note: This setting determines how many measurement points are actually taken every 3 min. Therefore, the output is the mean value of 11 measurement points over 11 s. - Enter the names for the samples.
- Skip the next screen, as the angle was already calibrated before (Step 4.1).
Note: A test run for the Bluetooth connection before inoculating the main cultures is advised to ensure stable connections and appropriate location of the connected computer. Also, the batteries have to be charged.
- Click Start Measurement, name the experiment and enable all parameters needed to monitor.
- Measure the cell density of the ONC (Step 3.1) diluted in cultivation media (1:20) in a spectrophotometer at 600 nm wavelength. Inoculate 50 mL of the medium (recommended carbon source concentration 0.3-1%, different media possible as described in Step 1.) with the ONC to an OD600 of 0.05 by using the following calculation.
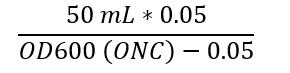
- Place the flasks in the detectors and shake at 28 °C, 130 rpm and 80% humidity.
Note: It is very important to give the culture 5 min of shaking before starting the measurement to avoid cells sticking to the bottom of the flask and yielding wrong values. - Click Start Measurement in the software.
- Take 0.2 mL samples for cell density measurement 4 h after the inoculation and when the carbon source gets depleted (determination described in step 4.7) in triplicates. Dilute the samples appropriately in cultivation media (first measurement 1:5, second measurement 1:20) and measure the absorption at 600 nm wavelength in a spectrophotometer. Before removing the flasks and even before stopping the shaker, always pause the measurement in the software.
Note: The detectors are calibrated for recording the parameters under defined shaking conditions and will yield to nonsense measurements when recording still cultures. Also, make sure to always wait 5 min of shaking before starting the measurement again. The cell density measurements are essential because the detector is delivering just values for the biomass and not real cell density. A calibration function between the OD600 values (x values) and the values obtained by the online measurement system (y values) can be achieved by measuring several timepoints spectrophotometrically. The values are not in a linear but in a logarithmic relation according to the following calibration function:
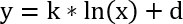
y: OD600 values obtained by spectrophotometer
x: values obtained by online measurement system
k: slope
d: axis intercept
The slope and the axis intercept can then be used to convert any value into the corresponding OD600 value. In Excel one can plot the spectrophotometrically obtained OD600 values against the values from the online system from the same timepoint. The addition of a logarithmic trendline and the corresponding equation will give the calibration function shown above. - Cultivate until the carbon source is nearly depleted (app. 12-20 h). For that purpose, let the cultures grow until it can be seen in the software diagrams that the oxygen concentration is approaching zero and the cells are in the exponential growth phase.
Note: This is the case for carbon source concentrations used in this protocol. The timepoint of carbon source depletion can be determined with the online monitoring system as it is shown exemplarily in Figure 1 in the Representative Results section, when the oxygen level approaches zero and the cells are in the exponential growth phase.
5. Protein Expression by De-repression
- When the timepoint described in Step 4.6 is reached, start feeding the cultures by the addition of four feed discs per flask under sterile conditions.
Note: Under the described conditions, four glycerol feed discs release 2 mg/h glycerol according to the manufacturer. - Continue the cultivation for 60-90 h while taking samples every 24 h to monitor the protein expression by an assay of choice (e.g., Bradford assay15, SDS PAGE16 or activity assays) and cell density measurements.
6. Culture Harvesting
- After the 60-90 h of cultivation, fill the cultures in 50 mL centrifuge tubes for harvesting by centrifugation at 4,000 x g, 4 °C for 10 min.
Note: The supernatant (for secreted proteins) or the cells (for intracellular protein expression) can be harvested by centrifugation at 4,000 x g, 4 °C for 10 min and further analyses can be done. - Export the online measurement data as a spreadsheet file from the software for further analysis.
- Fill 5-10 mL of distilled water to all the flasks and autoclave to inactivate the remaining cells.
As described in the protocol section, the software records the important cultivation parameters over the whole cultivation. Figure 1 shows the visualization of biomass development and oxygen concentration during a cultivation started with either 0.5 % (Figure 1A) or 0.3 % (Figure 1B) carbon source in the batch followed by the addition of glycerol feed discs. The online monitoring system is especially useful for the de-repressed cultivations as the timepoint of carbon source depletion after the batch can be determined exactly. As it can be seen in Figure 1, the oxygen concentration in the medium is decreasing rapidly and approaching zero, especially for 0.5 % glycerol in the batch (Figure 1A) while the biomass is increasing exponentially at this point. Here, the cells are in the exponential growth phase and in order to make optimal use of the carbon source release, shortly before the culture reaches the stationary phase, the feed discs should be added as it is described in the protocol. The addition of the feed discs before the oxygen concentration reaches zero is important for de-repressed protein expression to prevent the cells from oxygen limitation. Lower glycerol concentrations reduce the effect of short-term oxygen limitations as it can be seen in Figure 1B and are therefore recommended for de-repressed protein expression.
In the following, the results of two different cultivation experiments – employing the described protocol and the monitoring system shown in Figure 1 – are described. In the first experiment, a P. pastoris strain producing the glucose oxidase from Aspergillus niger (A. niger) under the control of the PDC and in the second a P. pastoris strain producing the human growth hormone under control of the PDF is shown.
In this experiment, glucose oxidase production under the control of PDC was investigated. Therefore, different cultivation strategies were compared: glycerol pulsing, constant glycerol feed by addition of feed discs and cultivation without any feed or pulsing. The cultivation was done in 50 mL buffered minimal media with 0.5% glucose as sole carbon source in 250 mL shake flasks (Table 1).
Upon 160 h cultivation, the highest biomass concentrations were obtained, when a feeding strategy using a glucose batch for biomass production (38 h) and glycerol pulsing (0.25% w/v glycerol after 38 h, 61 h and 86 h cultivation) was employed (0.21 g/L total secretory protein, 8.4 U/mL). Although a 2-fold reduction of the biomass concentration and the total amount of secreted protein were obtained, when the feed disks were added after 38 h glucose batch, the volumetric activity was even 1 U/mL higher compared to the cultivation using glycerol pulsing. This indicates that a substantial amount of secreted protein does not contribute to the volumetric activity in the supernatant and may be secreted in an inactive form. Therefore, higher glycerol concentrations from glycerol pulses seemingly favor the formation of biomass instead of target protein production and constant low glycerol release led to more than two-fold higher specific productivity.
Another experiment employing the de-repressed expression method was done by expressing the human growth hormone (hGH) under control of PDF. The construct was stably integrated into the genome of P. pastoris strain BSYBG11. Here, 50 mL of BYPG, buffered rich medium, in 250 mL baffled shake flasks was used with 0.3% (w/v) glycerol as carbon source in the batch. Upon glycerol depletion, the protein expression with constant glycerol feed was ensured by the addition of 4 feed discs per flask and compared to no glycerol feed (Figure 2).
Figure 2 shows the comparison between hGH expression and biomass development of the same strain cultivated with and without constant glycerol feed. As assay sandwich ELISA was used with the secondary antibody fused to HRP and detection was carried out with 3,3′,5,5′-tetramethylbenzidine (TMB) as substrate. Especially for this protein, it seemed to be the case that without any feeding, the cells start to degrade the produced protein after approximately 30 h (light blue squares), whereas by feeding, this degradation could be prevented (dark blue triangles) and even the protein concentration per biomass still increased after 150 h of cultivation. As already observed for Hansenula polymorpha14, this result shows how switching from batch, like it is usually used in shake flask scale, to fed-batch mode can optimize P. pastoris cultivations.
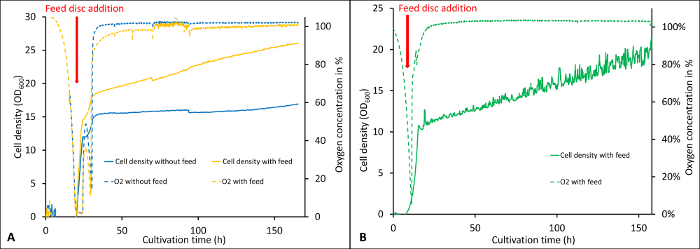
Figure 1: Visualization of online recorded parameters during two cultivations with different glycerol concentrations in the batch. The cultivations were started with 0.5% w/v (A) and 0.3% w/v (B) glycerol in the batch followed by the de-repression phase where the cells were fed by applying 4 feed discs per flask (release rate of 0.5 mg/h/disc according to the manufacturer). The dashed lines show the oxygen concentration in the medium, whereas the continuous lines show the cell density (OD600). Error bars show the standard deviation of six replicates (2 biological, three technical each). Please click here to view a larger version of this figure.
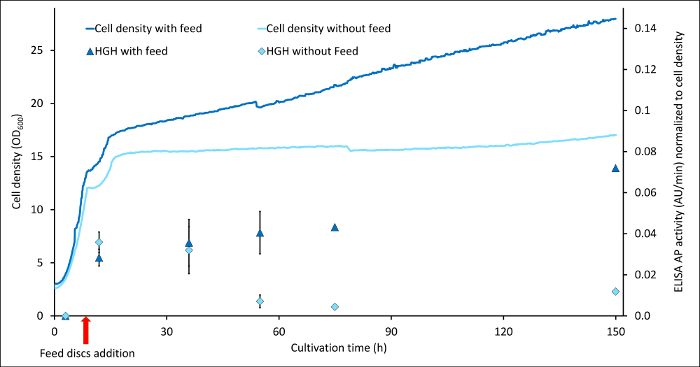
Figure 2: Comparison of human growth hormone (hGH) expression in batch and fed-batch mode. hGH content is indicated by absorption values at 405 nm by an ELISA where the 2nd antibody was an HRP fusion and TMB was used as a substrate for detection. Shown are the results for two different cultures: light blue line and squares display the cell density and hGH concentration normalized by the cell densities of a culture without any feed after the batch, compared to a culture, where the glycerol feed discs were applied (dark blue lines and triangles). Error bars show the standard deviation of six replicates (2 biological, three technical each). Please click here to view a larger version of this figure.
| PDC-GOX at 160 h cultivation time | OD600 | vol. activity [U/mL] | total amount of secreted protein [mg/mL] | volumetric activity per cell density | |||
| Mean | SD | Mean | SD | Mean | SD | Mean | |
| BMD1% batch + glycerol pulsing | 74.6 | 2 | 8.4 | 2.4 | 0.21 | 0.01 | 0.11 |
| BMD1% batch + constant glycerol feed | 32.5 | 0.3 | 9.4 | 0.7 | 0.09 | 0.01 | 0.29 |
| BMD1% batch | 18.8 | 0.6 | 3.7 | 0.7 | 0.01 | 0 | 0.2 |
Table 1: Results of methanol independent GOX expression under the control of the de-repressed promoter PDC.
