Morphology of DMSO treated iPSCs
Human iPSCs derived from control subjects were cultured either in an adherent 2D monolayer or in 3D cell spheres in suspension. Approximately 24 h after initial plating, cells were treated with either 1% or 2% DMSO for 24 h in the maintenance medium. Representative brightfield images after DMSO treatment are shown in Figure 1. Consistent with previous reports for iPSCs maintained in a monolayer3, DMSO pretreatment resulted in a transient dose-dependent decrease in growth rate as compared to non-DMSO treated cells (Figure 1A). This decreased proliferation is associated with an increase in cell-to-cell contact, which is especially pronounced in the 2% DMSO treated cells displaying increased formation of more highly clustered cell colonies. In other cell types, DMSO-induced G1 arrest has been shown to be associated with increased expression of proteins involved in cell-cell interactions that support contact-inhibition induced growth arrest36. In iPSCs maintained as 3D cell spheres, the DMSO treatment similarly increased the number of cell spheres (Figure 1B). Furthermore, DMSO treatment also resulted in less variable 3D sphere sizes, which has been previously shown to be indicative of improved differentiation capacity of the cells37. Importantly, neither 1% or 2% DMSO resulted in cell toxicity, as measured by viability counts (n = 3; 2D culture % live = control: 80 ± 1.3; 1% DMSO: 82 ± 3.7, 2%: 81 ± 2.7; 3D culture % live = control: 81 ± 4.3; 1% DMSO: 82 ± 6.7, 2%: 82 ± 2.7). Overall, these results are consistent with the notion that DMSO treatment alters the cell cycle and growth patterns in cultured stem cells. These effects on growth inhibition are reversible when the DMSO is removed from the medium, as previously shown3.
DMSO treatment improves the differentiation of ESCs to the primary germ layers
HUES6 hESCs were seeded on coated plates for 24 h followed by treatment with 2% DMSO for 24 h in the maintenance medium. Cells were then differentiated into the three primary germ layers following the treatment paradigms shown in Figure 2A30,31,32. Differentiated cells were then fixed and immunologically stained for prototypic markers of each respective germ layer (SOX17 for endoderm, brachyury for mesoderm, and SOX1 for ectoderm). As shown in Figure 2B, 24 h of pretreatment with 2% DMSO increased the proportion of cells expressing each respective germ layer marker. This is consistent with previous reports from our lab showing increased immunoreactivity, gene expression, as well as absolute number of differentiated cells towards all germ layers in stem cells treated with DMSO3,5. HUES6 is an hESC line with very low propensity for differentiation across all lineages1, yet the DMSO treatment substantially improves its capacity to differentiate across all germ layers.
DMSO treatment improves the differentiation to progenitor cell types
To investigate the effect of DMSO on differentiation to CNS progenitor cell types, human iPSCs were differentiated to either neural progenitor cells (NPCs) or oligodendrocyte progenitor cells (OPCs). To generate NPCs, cells were pretreated with 2% DMSO for 24 h in the maintenance medium followed by 12 days of directed differentiation33 (Figure 3A). As shown in Figure 3B, 2% DMSO pretreatment increased the expression of the NPC marker PAX6 as compared to control. Using another previously validated protocol34 (Figure 3C), iPSCs were differentiated for 12 days into OPCs. Similar to NPCs, OPCs derived from iPSCs pretreated with 2% DMSO for 24 h demonstrated an increase proportion of cells expressing OPC markers OLIG2 (Figure 3D).
An initial DMSO treatment persists to enhance differentiation into mature cell types
To investigate the effect of DMSO on latter stages of a differentiation protocol, HUES8 hESCs were pretreated for 24 h with 2% DMSO prior to differentiation to β cells following a 20 day directed differentiation protocol described in Figure 4a35. HUES8 were used as they have been previously shown to have a higher propensity towards endodermal lineage1,38. At the definite endoderm stage, the differentiated cells express SOX17 and FOXA2, definitive endoderm (DE) specific markers. With further differentiation into the pancreatic progenitors (PP1) stage, differentiated cells express PDX1 and FOXA2, markers characteristic of pancreatic progenitor cells. At these stages of pancreatic cell differentiation, the efficiencies of induction into DE and subsequently into PP1 were high for both control and DMSO-treated hESCs differentiated into each of these stages (Figure 4B, stages 1 and 3). Even though the HUES8 cell line has been noted to have increased propensity to differentiate into the endodermal lineage, as differentiation is induced further into the more specialized cell types at the terminal stages the DMSO-treated hESCs are much more likely to produce mature pancreatic endocrine cells. The efficiencies of generating PDX1/NKX6.1+ pancreatic progenitor cells, Neurogenin 3+ endocrine cells, and NKX6.1/C-peptide+ SC-β cells were substantially higher in the DMSO-treated hESCs (Figure 4B, stages 4 and 5). These results are in line with the NPC and OPC differentiation showing that DMSO enhances the differentiation potential to progenitor cell types and also demonstrates that the effect of DMSO is persistent in generating more specialized cell types. This is consistent with prior work, where we have shown that the initial 24 h DMSO treatment increases differentiation into terminal cell types across germ layers, including into neuronal cells as well as beating cardiomyocytes31,39 in cell lines with high or poor propensities for differentiation3.
Initial DMSO treatment improves hESC-derived cell function following in vivo transplantation
Previously, we have demonstrated the effectiveness of DMSO treatment in enhancing the differentiation of hESCs into functional pancreatic progenitor cells that later show a marked improvement in insulin secretion in vivo3. Using previously published protocols3,30,40, HUES8 hESCs were treated with 1% DMSO for 24 h, differentiated into pancreatic progenitor cells, and transplanted into immunodeficient SCID-Beige mice to assess functionality (e.g., insulin secretion in response to a glucose challenge or KCl stimulation) (Figure 5A). While the efficiencies of differentiation into FOXA2+ (~90%) and PDX1+ (~75%) pancreatic progenitors were comparable between control and DMSO-treated hESCs (Figure 5B) for the HUES8 hESC line, the cells differentiated from hESCs following a 24 h 1% DMSO treatment had improved responsiveness to glucose and KCl stimulation following in vivo transplantation. Improvements in functionality were evident within 2 weeks post-transplantation (Figure 5C) and persisted up to at least 16 weeks post-transplantation (Figure 5D). Taken together, these results suggest that DMSO pretreatment not only increases the differentiation efficiency to germ layers, progenitor cells, and more mature cell types, but also that it persists to enhance functionality of the differentiated cells in vivo.
| Differentiated Cell Type | Starting Cell Type | %DMSO | Length of DMSO Treatment | Length of DMSO Treatment |
| Hepatic cells | ESC Hepatoma cell line ESC ESC Mesenchymal stem cells iPSCs ESC ESC Hepatoma cell line ESC |
1.0 1.0 1.0 0.5 0.1-2.0 1.0 1.0 0.5 1.0 0.6 |
8 days Several days 7 days 10-14 days 7-21 days 7 days 4 days 5 days 2-21 days Throughout |
Basma et al., 2008 Kanebratt and Andersson, 2008 Hay et al., 2009 Duan et al., 2010 Alizadeh et al., 2014 Kondo et al., 2014 Szkolnicka et al., 2014 Czysz et al., 2015 Nikolaou et al., 2016 Vanhove et al., 2016 |
| Primary germ layers | ESCs and iPSCs hESC hESC |
0.1-2.0 0.5 0.1-2.0 |
24 hours 24 hours 24 hours |
Chetty et al., 2013 Chetty et al., 2015 Li et al., 2018 |
| Cardiac cells | ESCs and iPSCs P19 cells ESCs and iPSCs Fetal mesenchymal stem cells |
0.1-2.0 1.0 1.0-2.0 0.8-1.0 |
24 hours 4 days 24-30 hours 24 hours |
Chetty et al., 2013 Choi et al., 2014 van den Berg et al., 2016 Deng et al., 2017 |
| Pancreatic cells | ESCs and iPSCs hESC |
0.1-2.0 0.5 |
24 hours 24 hours |
Chetty et al., 2013 Chetty et al., 2015 |
| Smooth muscle cells | P19 cells | 1.0 | 4 days | Choi et al., 2014 |
| Endothelial cells | P19 cells | 1.0 | 4 days | Choi et al., 2014 |
| Enterocytes | iPSCs | 0-1.6 | 4 days | Ogaki et al., 2015 |
| Gut epithelium | iPSCs | 0-1.6 | 4 days | Ogaki et al., 2015 |
| Neural cells | Marmoset iPSC | 0.05-2.0 | 24 hours | Qiu et al., 2015 |
| Neutrophils | Leukemia cell line | 1.25 | 6-8 days | Teimourian and Moghanloo, 2016 |
| Skeletal Myotubes | iPSCs | 1.5 | 24 hours | Swartz et al., 2016 |
| Cortical organoid | hiPSCs | 1.0 | 24 hours | Yoon et al., 2018 |
Table 1: Summary of previously published work demonstrating the beneficial effects of DMSO treatment on differentiation.
| S1 | S2 | S3 | S5 | |
| MCDB131 (L) | 1 | 1 | 1 | 1 |
| Glucose (g) | 0.44 | 0.44 | 0.44 | 3.6 |
| NaHCO3 (g) | 2.46 | 1.23 | 1.23 | 1.754 |
| FAF-BSA (g) | 20 | 20 | 20 | 20 |
| ITS-X (mL) | 0.02 | 0.02 | 5 | 5 |
| Glutamax (mL) | 10 | 10 | 10 | 10 |
| Vitamin C (mg) | 44 | 44 | 44 | 44 |
| Heparin (mg) | 0 | 0 | 0 | 10 |
| P/S (mL) | 10 | 10 | 10 | 10 |
Table 2: Components of endocrine progenitor cell differentiation base medias.
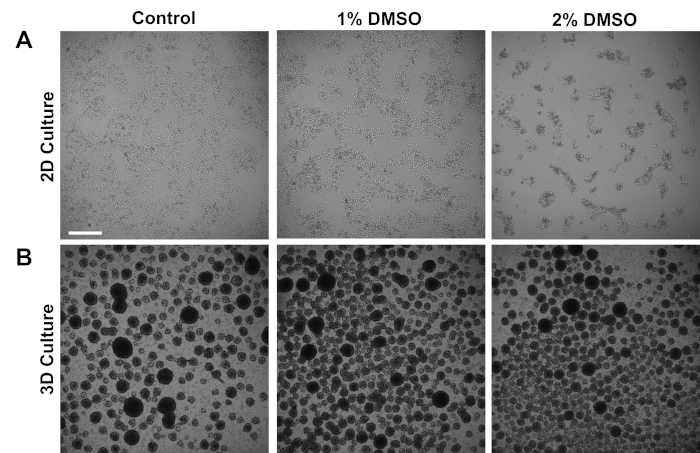
Figure 1: DMSO treatment alters the growth of hPSCs. (A) Representative brightfield images of hiPSCs plated in a monolayer after receiving no treatment (control) or treatment with 1% or 2% DMSO for 24 h. DMSO promotes a transient dose-dependent growth inhibition of iPSCs. (B) Representative brightfield images of hiPSCs plated on low-attachment plates to allow 3D sphere formation after receiving no treatment (control) or treatment with 1% or 2% DMSO for 24 h. DMSO treatment results in less variable 3D sphere formation compared to control. Scale bar = 500 µm. Please click here to view a larger version of this figure.
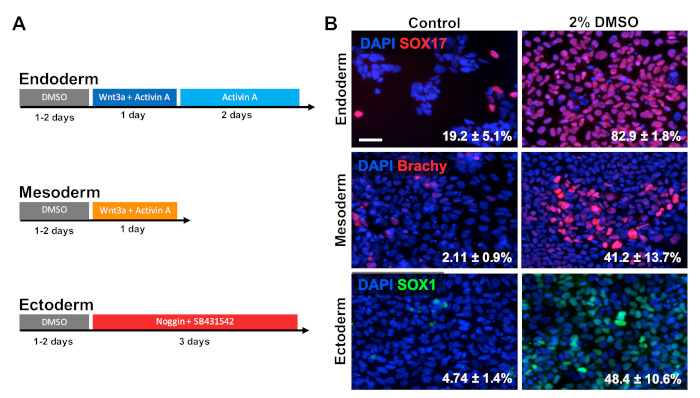
Figure 2: DMSO treatment improves differentiation of hPSCs to primary germ layers. (A) Schematic of differentiation protocols used to generate the three primary germ layers. (B) Representative images of differentiated HUES6 hESCs immunolabeled for SOX17 (endoderm), brachyury (mesoderm), and SOX1 (ectoderm). Pretreatment with 2% DMSO for 24 h increased the differentiation efficiency across all three germ layers. Percentages of cells differentiating into SOX17+ endodermal, Brachyury (Brachy)+ mesodermal, or SOX1+ ectodermal cells following directed differentiation into each germ layer of control and DMSO-treated hESCs are noted with SEM of three biological replicates. Unpaired t-test: endoderm p = 0.0003; mesoderm p = 0.047; ectoderm p = 0.015. Scale bar = 50 µm. Please click here to view a larger version of this figure.
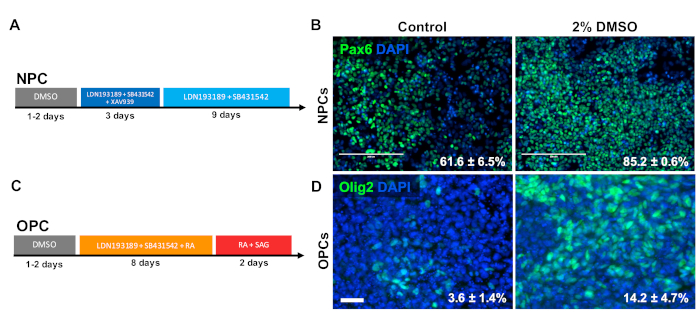
Figure 3: DMSO treatment improves differentiation to neural progenitor cell types. (A) Schematic of differentiation protocol used to generate neural progenitor cells (NPCs). (B) Representative images of human iPSCs differentiated into NPCs immunolabeled for Pax6. 24 h of pretreatment with 2% DMSO increased the number PAX6 positive cells. Percentages of cells differentiating into Pax6+ NPCs following directed differentiation of control and DMSO-treated human iPSCs are noted with SEM of three biological replicates. Unpaired t-test: p = 0.0225. Scale bar = 200 µm. (C) Schematic of differentiation protocol used to generate oligodendrocyte progenitor cells (OPCs). (D) Representative images of human iPSCs differentiated into OPCs immunolabeled for OPC markers Olig2. 24 h of pretreatment with 2% DMSO increased the expression of both OPC markers compared to control. Percentages of cells differentiating into Olig2+ OPCs following directed differentiation of control and DMSO-treated human iPSCs are noted with SEM of four biological replicates. Unpaired t-test: p = 0.0466. Scale bar = 50 µm. Please click here to view a larger version of this figure.
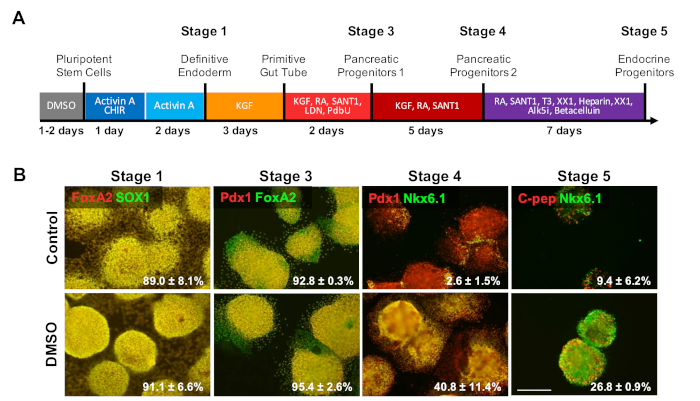
Figure 4: DMSO treatment enhances terminal differentiation potential of hPSCs. (A) Schematic of a ~20 day directed differentiation of HUES8 hESCs into terminally differentiated pancreatic endocrine cells. (B) Immunostaining for the indicated markers at each stage of differentiation following directed differentiation of untreated control cells and cells pretreated with 2% DMSO for 24 h. The initial DMSO treatment persists to increase differentiation into terminal endocrine cell types at the latter stages of directed differentiation. Percentages of cells differentiating into the indicated markers at each stage of differentiation following directed differentiation of control and DMSO-treated hESCs are noted with SEM of two to four biological replicates. Scale bar = 200 µm. Please click here to view a larger version of this figure.
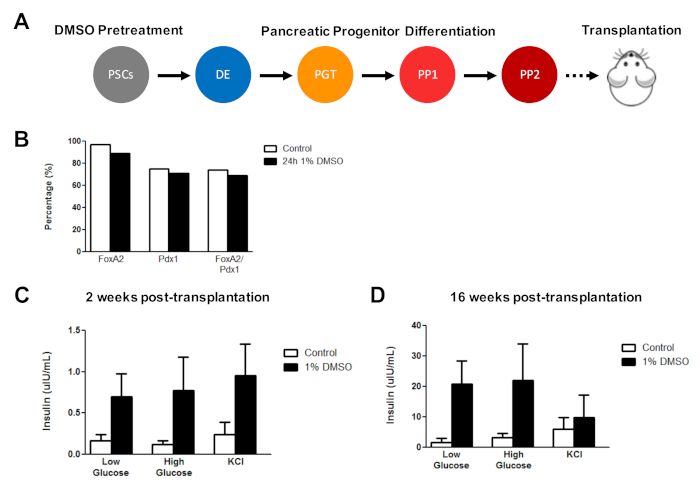
Figure 5: Initial DMSO treatment of hPSCs enhances glucose responsiveness following transplantation of pancreatic progenitor cells in vivo. (A) Schematic of directed differentiation (~15 days) of HUES8 hESCs into pancreatic progenitor cells (PP2) following no treatment (control) or a 24 h 1% DMSO treatment and subsequent transplantation (5 million cells) into immunodeficient SCID-Beige mice. (B) Percentage of cells differentiating into PDX1+ and FOXA2+ pancreatic progenitor cells following in vitro directed differentiation of control and DMSO-treated hESCs immediately before transplantation (n = 1). (C) Mean ELISA measurements of human insulin from the serum of mice following a low (2.5 mM) or high (15 mM) glucose challenge or potassium chloride (KCl) stimulation at (C) 2 weeks and (D) 16 weeks post-transplantation of pancreatic progenitor cells differentiated from control and DMSO-treated hESCs (error bars = SEM; n = 3 at 2 weeks and 16 weeks for control; n = 2 at 2 weeks and 16 weeks for DMSO). Two-way ANOVA: p = 0.0051 for control vs. DMSO at 2 weeks; p = 0.0116 for control vs. DMSO at 16 weeks. The mice studied at the different time points are different. Results are adapted from Chetty et al.3. Please click here to view a larger version of this figure.
