The protocol outlined in Figure 1 describes the basic workflow for utilizing the enzyme-coupled malachite green assay to probe the interaction of small molecules with the dNTPase SAMHD1 and can be adapted in a number of ways to interrogate different biochemical questions. In the representative results discussed in the below paragraphs, we illustrate examples of using this assay to determine the inhibitory properties of small molecules toward SAMHD1 and to test whether different nucleotide analogues are substrates and/or activators of this enzyme.
The results shown in Figure 2 illustrate several core principles of this assay. The malachite green reagent allows the colorimetric detection of inorganic phosphate through the formation of a phosphomolybdate malachite green complex, and accordingly, this approach can be applied to the study of enzymatic reactions whose product is phosphate. To demonstrate the sensitivity of this method to detect free inorganic phosphate, Figure 2A shows the absorbance values obtained with increasing concentrations of Na3PO4 following a 20 min incubation with the malachite green reagent. While the signal reaches saturation at 0.25 mM Na3PO4, the linear detection range of phosphate is visible from 0.004 to 0.03 mM (Figure 2A, right panel), in agreement with other studies that reported a phosphate linear range up to 10-20 μM using the malachite green assay38.
SAMHD1 is a dNTPase that releases inorganic triphosphate when hydrolyzing a dNTP molecule, and thus in order to generate free inorganic phosphate for detection by malachite green, a coupling enzyme is required. Inorganic pyrophosphatase (PPase) from E. coli has been shown useful for this purpose, both with regard to SAMHD17,20, but also other nucleotide metabolizing enzymes30,33,35. Additionally, SAMHD1 is an active dNTPase when as a homotetramer, and this requires allosteric activation by (d)NTPs, specifically a guanine triphosphate (GTP or dGTP) at AS1 and any dNTP at AS2. Subsequently, the catalytic site becomes accessible for substrate binding and the enzymatic reaction takes place. As dGTP fulfils the requirements for binding to AS1 and AS2, and is a substrate, use of this nucleotide in the inhibition assay greatly simplifies the workflow. Figure 2B illustrates the requirement of the different assay components to achieve measurable SAMHD1 activity indicated by an increase in absorbance at 630 nm. Neither SAMHD1 nor PPase alone are capable of generating inorganic phosphate in the presence of dGTP, consistent with the documented activities of these enzymes. However, in the condition in which all the assay components are present (SAMHD1, PPase, and the dGTP activator/substrate) we observe an increase in signal. The Z factor37 of the example shown here (taking no enzymes + dGTP as a negative control and SAMHD1/PPase + dGTP as a positive control) was 0.74, indicating a robust assay.
One of the potential applications of the enzyme-coupled SAMHD1 activity assay is the identification of inhibitors through high-throughput screening (HTS). Thus, in this report, we validate the detection of SAMHD1 inhibition in this assay using a diverse set of compounds already described in the literature. Seamon et al. evaluated the dose-dependent inhibition of canonical nucleosides toward SAMHD1 using a similar assay as shown here, and found that deoxyguanosine (dGuo) was the only canonical nucleoside able to significantly inhibit SAMHD1, with an IC50 value of 488 µM20. A HTS of FDA-approved drugs performed with the direct b4NPP assay revealed several hits that inhibited SAMHD1 activity at micromolar concentrations, from which lomofungin was the molecule that most potently inhibited SAMHD1 dNTPase activity in vitro, exhibiting an IC50 of 20.1 µM when determined in the presence of dGTP as a substrate21. Additionally, the four α,β-imido-dNTP analogs have also been identified as competitive inhibitors of SAMHD1 using the MDCC-PBP sensor and SAMHD1 coupled to Ppx activity, which showed that the inhibitory constants of the dNMPNPP analogs were in the low micromolar / high nanomolar range6,22. Thus, to demonstrate that the enzyme-coupled SAMHD1 activity assay can be used to identify SAMHD1 inhibitors, dGuo, lomofungin and 2'-deoxythymidine-5'-[(α,β)-imido]triphosphate (dTMPNPP), were used to validate the technique. Figure 3A illustrates the dose-response curves obtained for these compounds, showing that increasing concentrations effectively inhibit SAMHD1 activity. The mean IC50 values obtained for these molecules from three independent experiments (± standard deviation) were as follows: dGuo = 361.9 ± 72.8 µM, lomofungin 6.78 ± 3.9 µM, and dTMPNPP = 2.10 ± 0.9 µM. As an example of a negative result, the impact of hydroxyurea (HU) on SAMHD1 activity was also determined. HU is an inhibitor of ribonucleotide reductase, and, although it limits SAMHD1 ara-CTPase activity in various AML models, the effects of HU on SAMHD1 were shown to be indirect and rely on perturbing the allosteric regulation of SAMHD118. The dose response curve of HU is shown in Figure 3B, and no changes in SAMHD1 activity were observed with increasing HU doses, demonstrating that HU does not inhibit SAMHD1 activity in vitro.
Another use of the enzyme coupled SAMHD1 activity assay is to interrogate whether nucleotides and their analogs are substrates and/or allosteric activators of this enzyme, which is illustrated in Figure 4. In this experiment, canonical nucleotides, as well as the active metabolites of several anti-cancer nucleoside analogs, such as cytarabine (ara-CTP), clofarabine (Cl-F-ara-ATP), and gemcitabine (dF-dCTP), were tested as SAMHD1 substrates and activators. Due to the complex allosteric regulation of SAMHD1, the reaction is performed in the presence of GTP as an AS1 activator or the non-hydrolysable dGTP analog 2'-deoxyguanosine-5'-(α-thio)-triphosphate (dGTPαS), which can occupy AS1 and AS2. SAMHD1 activity in the presence of the tested nucleotide analog and GTP indicates that the nucleotide is able to bind to the secondary allosteric site and catalytic site (i.e., AS2 activator and substrate), while SAMHD1 activity with the nucleotide analog and dGTPαS indicates the nucleotide can only occupy the catalytic site (i.e., only a substrate). If the nucleotide is able to bind to both the AS1 and AS2 allosteric sites and to the catalytic site, SAMHD1 will be active in the presence of the nucleotide alone, as shown in the case of dGTP. The results show that all canonical dNTPs are able to bind to the AS2 site and to the catalytic site. In the case of nucleotide analogs, clofarabine triphosphate is an AS2 activator and a substrate, whereas cytarabine triphosphate is only able to occupy the catalytic site. On the other hand, no activity was observed with gemcitabine triphosphate, suggesting that under the conditions tested gemcitabine triphosphate is not able to act as allosteric effector nor substrate. Although this result is consistent with previous predictions9, later crystallization and kinetic studies10 revealed that gemcitabine triphosphate is able to bind the SAMHD1 catalytic pocket, and that it is indeed a substrate of the enzyme. However, in the latter study10, the authors show that the hydrolysis rate is considerably lower compared to other reported substrates, such as cytarabine triphosphate, thus explaining why we were not able to observe this with this screening setup.
Altogether, these representative results validate the use of the enzyme coupled SAMHD1 activity assay as a robust technique for the identification and characterization of SAMHD1 inhibitors, allosteric regulators, and substrates. However, similar to all experimental approaches, this method has its caveats, and so orthogonal assays (e.g., using a different assay technology) should be used to further validate findings.
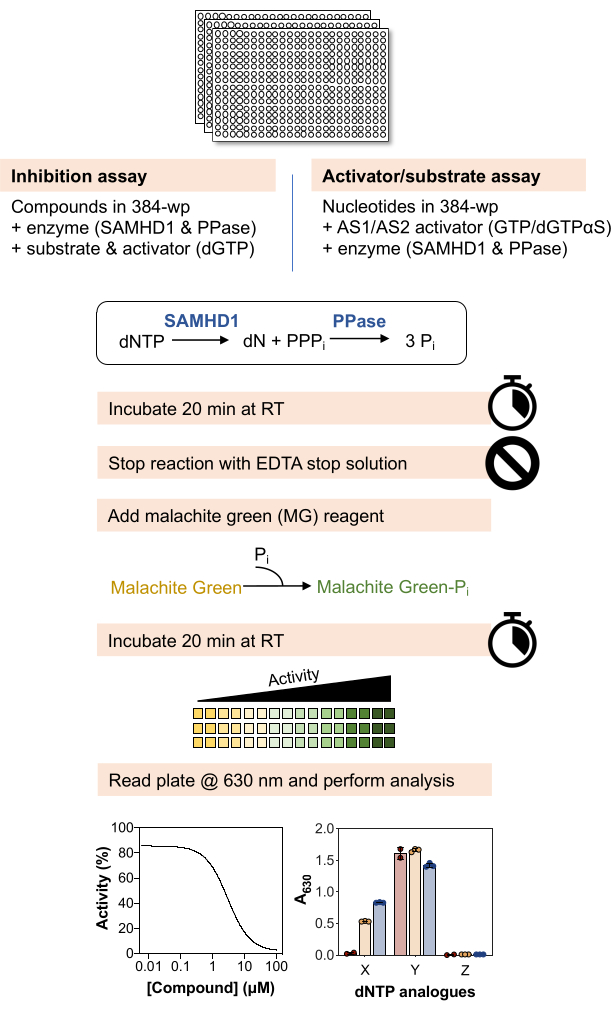
Figure 1: Schematic overview of the protocol described in this article. Please click here to view a larger version of this figure.
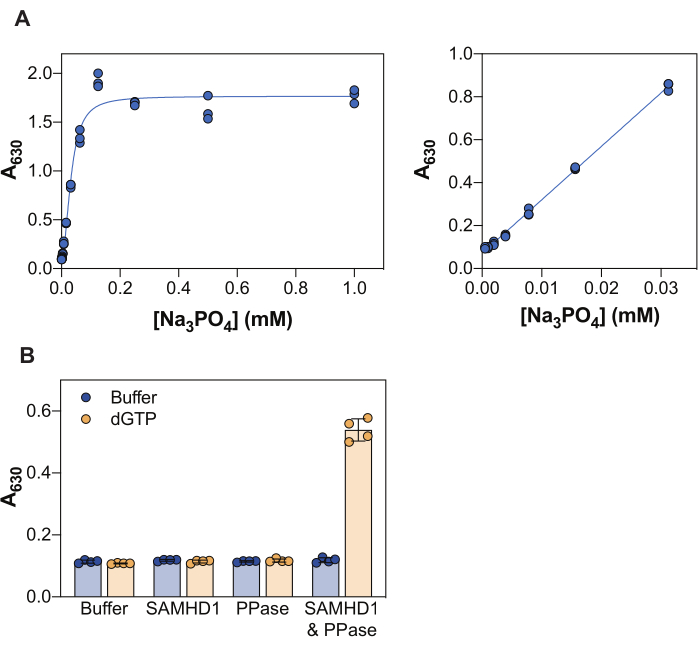
Figure 2: Enzyme-coupled SAMHD1 activity assay. (A) Na3PO4 standard curve in the malachite green assay. Na3PO4 serial dilution (2-fold) was prepared from 1 mM to 0.004 mM in triplicate and incubated with malachite green reagent for 20 min. Raw absorbance values over the full range of tested concentrations are shown in the left panel and the linear range in the right panel. Representative of two independent experiments shown. (B) Validation of enzyme-coupled activity assay. SAMHD1 (0.35 µM) and/or PPase (12.5 U/mL) in the presence or absence of activator/substrate dGTP (25 µM) were incubated for 20 min in the enzyme-coupled activity assay. Quadruplets from a representative of two independent experiments shown with raw absorbance values plotted, bars, and error bars indicate mean and SD. Please click here to view a larger version of this figure.
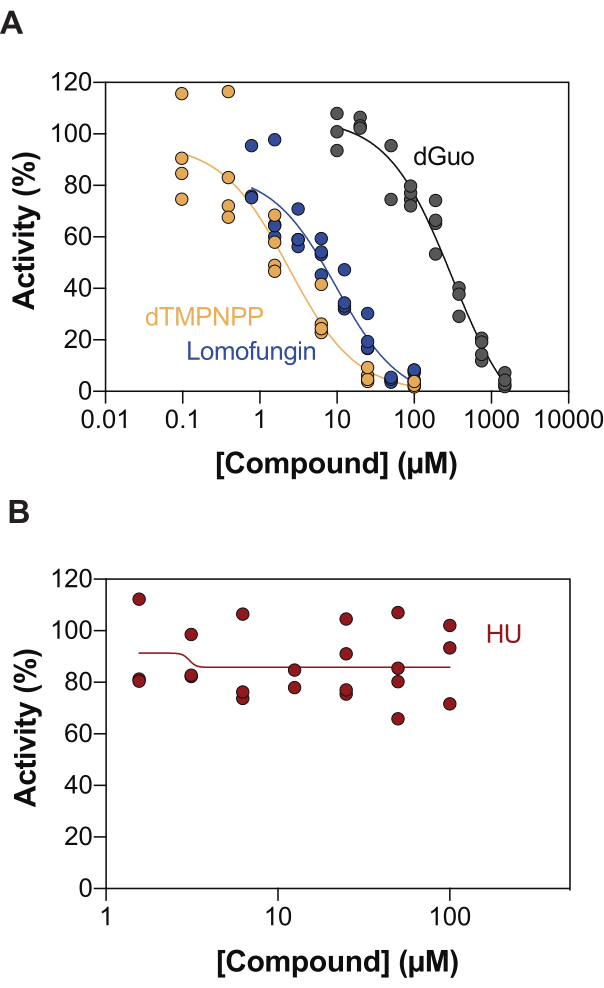
Figure 3: Evaluating compounds for SAMHD1 inhibition in the enzyme-coupled activity assay. Dose response of lomofungin (0.78-100 µM), 2'-deoxythymidine-5'-[(α,β)-imido]triphosphate (dTMPNPP, 0.01-100 µM) and deoxyguanosine (dGuo, 10-1,500 µM) (A) or hydroxyurea (HU) (0.78-100 µM) (B) in the enzyme-coupled SAMHD1 activity assay with dGTP (25 µM) as activator/substrate. Percentage activity relative to reaction controls from individual replicates plotted (DMSO + SAMHD1/PPase + dGTP = 100% activity, DMSO + dGTP = 0% activity) with a representative of three experiments shown. Please click here to view a larger version of this figure.
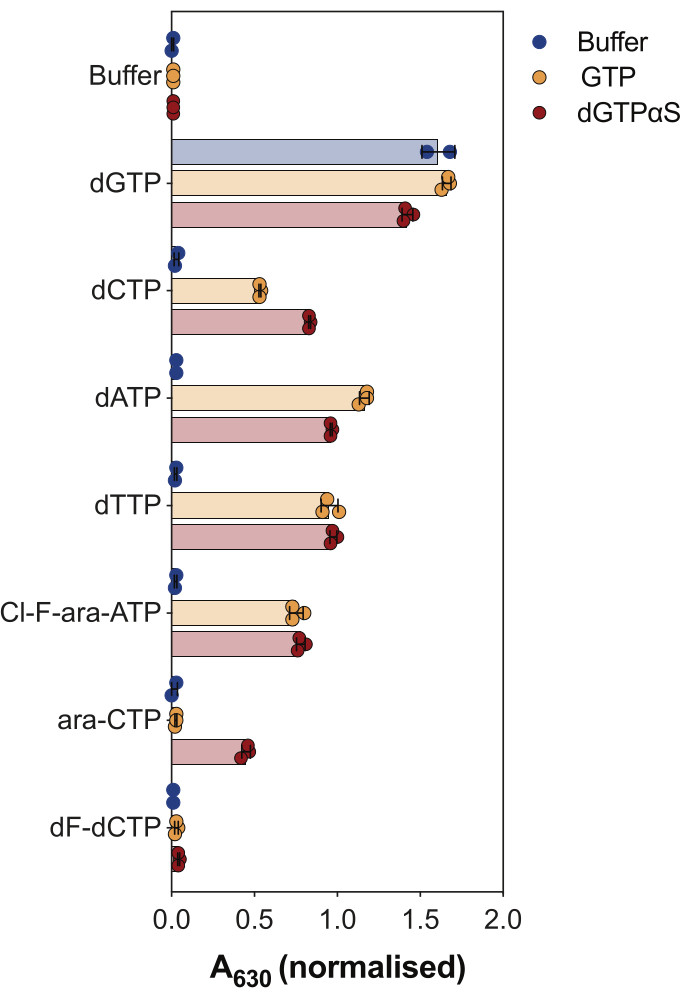
Figure 4: Evaluating nucleotide analogs as SAMHD1 allosteric activators and substrates in the enzyme-coupled activity assay. Canonical nucleotides and selected triphosphate metabolites of anticancer drugs cytarabine (ara-CTP), clofarabine (Cl-F-ara-ATP), and gemcitabine (dF-dCTP), were tested at 200 µM in the enzyme-coupled SAMHD1 activity assay in the presence or absence of GTP or non-hydrolysable dGTP analog dGTPαS (12.5 µM). Normalized absorbance values from individual experimental replicates plotted, mean and SD are indicated. Representative of two independent experiments shown, adapted from our previous study7. Please click here to view a larger version of this figure.
| Step | Reagent | Volume dispensed (µL) | Final reaction volume (µL) | Concentration dispensed | Fold dilution in reaction | Final concentration in reaction |
| 1 | Inhibitor | 5 | 20 | 0.4 mM | 4 | 0.1 mM |
| 2 | SAMHD1+PPase mix | 5 | 1.4 µM SAMHD1, 50 U/mL PPase | 4 | 0.35 µM SAMHD1, 12.5 U/mL Ppase | |
| 3 | dGTP | 10 | 50 µM | 2 | 25 µM | |
| 4 | Incubation for 20 minutes | |||||
| 5 | EDTA solution | 20 | 40 | 7.9 mM | 2 | 3.95 mM |
| 6 | MG reagent | 10 | 50 | 2.5 mM Malachite green, 64.4 mM Ammonium Molybdate, 0.18% Tween-20 | 5 | 0.5 mM Malachite green, 12.9 mM Ammonium Molybdate, 0.036% Tween-20 |
| 7 | Incubation for 20 minutes | |||||
| 8 | Read @ 630 nm | |||||
Table 1: Summary of the final conditions in the enzyme-coupled assay for inhibitors screening.
| Step | Reagent | Volume dispensed (µL) | Final reaction volume (µL) | Concentration dispensed | Fold dilution in reaction | Final concentration in reaction |
| 1 | Allosteric regulator | 5 | 20 | 800 µM | 4 | 200 µM |
| 2 | GTP or dGTPαS | 5 | 50 µM | 4 | 12.5 µM | |
| 3 | SAMHD1 and/or PPase | 10 | 0.7 µM SAMHD1, 25 U/mL PPase | 2 | 0.35 µM SAMHD1, 12.5 U/mL PPase | |
| 4 | Incubation for 20 minutes | |||||
| 5 | EDTA solution | 20 | 40 | 7.9 mM | 2 | 3.95 mM |
| 6 | MG reagent | 10 | 50 | 2.5 mM Malachite green, 64.4 mM Ammonium Molybdate, 0.18% Tween-20 | 5 | 0.5 mM Malachite green, 12.9 mM Ammonium Molybdate, 0.036% Tween-20 |
| 7 | Incubation for 20 minutes | |||||
| 8 | Read @ 630 nm | |||||
Table 2: Summary of the final conditions in the enzyme-coupled assay for allosteric regulators screening
