Ex vivo ERG enables recording of reproducible and stable photoreceptor and ON-bipolar cell light responses, for example, from the mouse retina (Figure 2A–C). Recording of photoreceptor responses from human donor retinas is possible with up to 5 h postmortem delay of enucleation (Figure 2D) and of ON-bipolar cell responses with a <20 min enucleation delay (Figure 2E). Important parameters to obtain large responses include a careful dissection technique, high perfusion rate, and perfusion temperature close to physiological values (35-38 °C in the mammalian retina). Under these conditions, response amplitudes and kinetics in both cell types were relatively stable over time but slowly declined approximately 40-45 min after retinas were mounted on the specimen holder (Figure 3).
Compared to photoreceptors, ON-bipolar cell function is more easily disrupted, for example, by damage to the retina during dissection and mounting or by a drop in the temperature and/or perfusion speed. While reduced temperature in the specimen holder greatly slowed the kinetics of both photoreceptors and ON-bipolar cells, it decreased the amplitude of the b-wave but not the a-wave (Figure 4A). Conversely, slowing the perfusion rate from 2.1 mL/min to 0.6 mL/min reduced the amplitudes of both photoreceptor and ON-bipolar cell responses but did not affect the implicit time (time from stimulus onset to response peak) of either the a- or the b-wave (Figure 4B). Cessation of perfusion for 10 min followed by reperfusion resulted in a complete loss of ON-bipolar cell function with preserved photoreceptor responses (Figure 5).
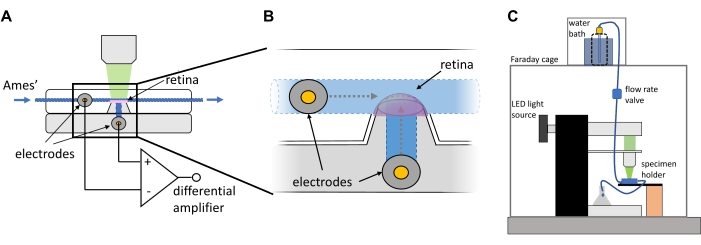
Figure 1: Ex vivo electroretinogram specimen holder and recording setup. (A,B) The ex vivo ERG specimen holder comprises a dome to mount the isolated retina, which is connected to a perfusion line to continuously deliver Ames' medium. Electrodes are connected through narrow channels to both the photoreceptor side of the retina via the perfusion line and the inner retina through the filter paper glued to the dome. These electrodes are connected to a differential amplifier, which enables measurement of potential differences in the retina in response to light stimuli. (C) The specimen holder is placed onto the stage of a microscope, which has been modified to deliver light flashes and connected to the perfusion line, which delivers heated, oxygenated Ames' medium by gravity. The entire recording setup is shielded by a Faraday cage to minimize electrical noise. This figure has been modified from 9. Abbreviation: ERG = electroretinogram. Please click here to view a larger version of this figure.
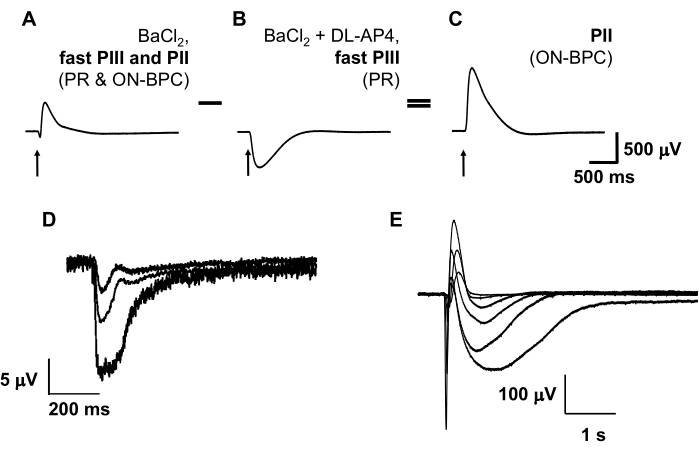
Figure 2: Example traces of ex vivo photoreceptor and ON-bipolar cell responses. Addition of pharmacological agents to the perfusate allows quantification of contributions from individual retinal cell types to the ex vivo electroretinogram. Photoreceptor (PR) light responses are isolated in the presence of 100 µM barium chloride (BaCl2), a blocker of K+ channels expressed by Müller glial cells, as well as 40 µM DL-AP4, a glutamate receptor blocker, which inhibits signal transmission from photoreceptors to ON-bipolar cells (B). ON-bipolar cell (ON-BPC) function (C) is determined by subtracting the photoreceptor component (B) from the combined photoreceptor and ON-bipolar cell response in the presence of barium chloride alone (A). Photoreceptor light responses can be obtained from human donor retinas with a death to enucleation delay of <5 h (D), whereas retinas enucleated within 20 min of death frequently also give ON-bipolar cell responses (E) (see 8 for further information). Figure 2A–C is modified from 9. Abbreviations: PR = photoreceptor; ON-BPC = ON-bipolar cell. Please click here to view a larger version of this figure.
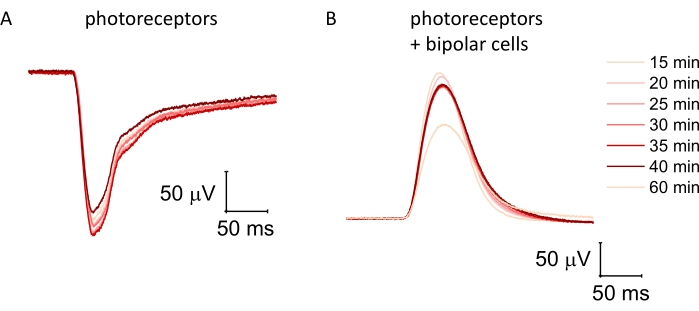
Figure 3: Stability of photoreceptor and ON-bipolar cell function in the ex vivo electroretinogram over time. (A) Light responses were recorded every minute from photoreceptors alone in the presence of both 100 µM barium chloride and 40 µM DL-AP4. (B) Dim light flashes in the presence of 100 µM barium chloride alone are heavily dominated by ON-bipolar cell function, although they contain a small photoreceptor component. Light responses from isolated retinas typically stabilize after perfusing the ex vivo specimen holder for 15-20 min, and were stable for at least 20-25 min before starting to decline. Please click here to view a larger version of this figure.
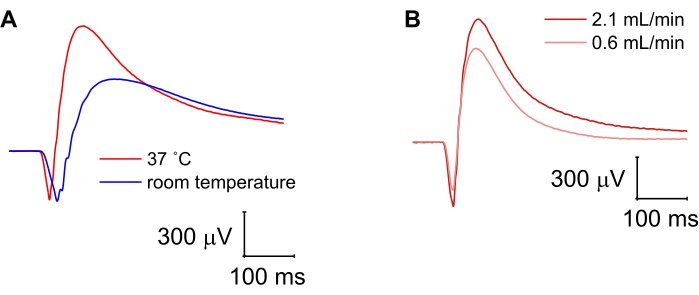
Figure 4: Combined photoreceptor and ON-bipolar cell responses at different temperatures and perfusion speeds. (A) Lowering of the temperature inside the specimen holder from 37 °C to room temperature greatly slowed photoreceptor and ON-bipolar cell kinetics but only reduced the ON-bipolar cell amplitude in the mixed photoreceptor and ON-bipolar cell response. (B) Reduction of the perfusion rate from 2.1 mL/min to 0.6 mL/min resulted in decreased photoreceptor and ON-bipolar cell amplitudes but no change in the response kinetics. Please click here to view a larger version of this figure.
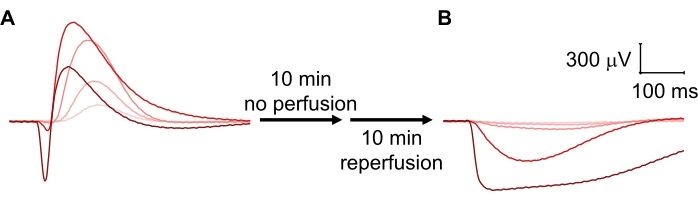
Figure 5: ON-bipolar cell responses are more sensitive to cessation of perfusion. (A) Large photoreceptor and ON-bipolar cell responses were recorded following perfusion with 2.1 mL/min for 20 min. (B) After cessation of perfusion for 10 min followed by reperfusion for 10 min at 2.1 mL/min, photoreceptor responses were present, whereas ON-bipolar cells responses were completely lost. Please click here to view a larger version of this figure.
