1. Cell preparation
- Perform all the experiments under sterile conditions in Class II laminar flow microbiological safety cabinet (see Table of Materials).
- Turn on the hood and allow for the stabilization of the airflow.
- Thoroughly spray the interior hood surface with 70% ethanol to eliminate any possible contamination from previous users.
- Prepare 5% of a disinfectant solution in a 500 mL glass beaker. Discard any cell supernatant or cell debris inside the solution.
- Thoroughly clean all the micropipettes and tip boxes with 70% ethanol.
- Prepare fresh cell culture media by supplementing Dulbecco's modified Eagle's medium (DMEM) high glucose media with 10% heat deactivated fetal bovine serum (FBS), 1% penicillin/streptomycin (100 unit/mL of penicillin and 100 µg/mL of streptomycin), and 2 mM L-glutamine. For the LX2 hepatic stellate cell line, reduce the concentration of the FBS supplement to 2%.
- Warm culture media before initiating the experiment.
- Take Huh7 and Hep3B HCC tumor cell lines and COS7 and LX2 fibroblast cell lines from their storage rack in liquid nitrogen. Rapidly defrost cryopreserved cells.
- Dilute thawed cells with 2 mL of fresh culture media. Centrifuge cells at 200 x g for 4 min at room temperature. Discard the supernatant and resuspend the cell pellet in 1 mL of fresh warm culture media.
- Seed cells in T75 cell culture flask. Incubate the cells in a cell culture incubator in 5% CO2 at 37 °C in 95% humidified conditions until cells reach 60%-70% confluency.
2. Cell collection
- Aspirate culture media and wash cells three times with phosphate buffer saline (PBS).
- Add 2 mL of pre-warmed 1x Trypsin to detach adherent cells from the bottom of the T75 flasks. Incubate at 37 °C in an incubator for 4 min.
- Inactivate trypsin by adding 4 mL of complete culture media. Collect the cell suspension and centrifuge cells at 200 x g for 4 min at room temperature. Discard the supernatant and resuspend the cells in 4 mL fresh culture media.
3. Cell counting
- Gently vortex the cell suspension to ensure homogenous distribution of cells in the centrifuge tube.
- Using a 10 µL pipette, mix 10 µL of cell suspension with 10 µL of Trypan blue. Gently pipette the mixture up and down four times to ensure complete staining of the outer cell surface with the dye.
- Count the number of cells using a hemocytometer.
- First, place a coverslip over the hemocytometer counting area before loading the stained cell suspension.
- Place the pipette tip containing the cell suspension into the V groove of the hemocytometer. Gently expel the tip content into the counting slide.
- Leave the slurry to settle for a couple of minutes before fixing it on the microscope stage for cell counting.
NOTE: To avoid double-counting, only count the cells on the two sides of the large square. - Count in cells overlapping the top or the right ruling and avoid those overlapping the bottom or the left ruling.
- Calculate the total number of cells.
NOTE: Cell number per ml =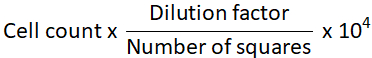
4. Collection of fibroblast conditioned media (CM)
- Aspirate culture media and wash the LX2 cells three times with PBS.
- Add 2 mL of pre-warmed 1x Trypsin to detach adherent cells from the bottom of the T75 flasks. Incubate the flask at 37 °C in an incubator for 4 min.
- Inactivate trypsin by adding 4 mL of complete culture media. Collect the cell suspension and centrifuge cells at 200 x g for 4 min at room temperature. Count the cells as per step 3.
- Seed 1 x 106 LX2 cells in 10 cm3 dishes for 48 h at 37 °C.
- Collect the fibroblast CM after 48 h. Centrifuge at 200 x g for 4 min at room temperature to pellet any floating cells. Sterile filter the CM using a 0.22 µm filter fixed on the bottom of 20 mL syringes.
- Collect the supernatant CM. Aliquot the CM into 2 mL tubes and store at -80 °C for further applications.
NOTE: The solution can be stored for 6 months at -80 °C.
5. Validating cell densities for perfect spheroids
- Aspirate culture media and wash the HCC cell lines three times with PBS.
- Add 2 mL of pre-warmed 1x Trypsin to detach adherent cells from the bottom of the T75 flasks. Incubate at 37 °C in an incubator for 4 min.
- Inactivate trypsin by adding 4 mL of complete culture media. Collect the cell suspension and centrifuge cells at 200 x g for 4 min at room temperature.
- Count the cells as per step 3.
- Pipette different densities from the tumor cell lines (12000, 6000, 3000, 1500, 1000, 750, 500, 250, and 125 cells) in 20 µL of media on the interior surface of a 10 cm3 lid of a Petri dish.
- Add 10 mL of sterile PBS to the bottom of the dish to provide humid conditions for the process of spheroid formation.
- Invert the lid of the 10 cm3 dish to allow the media, including the cell suspension, to hang over a humid environment. Leave the hanging droplets for 3 days.
- Take images of the spheroids at 50x magnification using an inverted microscope 3 days after hanging the original droplets.
6. Heterotypic tumor/stromal spheroids
- Suspend 1500 Huh7 HCC cells with 1500 COS7 mammalian fibroblast cells (1:1 ratio) in hanging droplets to form spheres. Add 10 mL of sterile PBS to the bottom of the dish to provide humid conditions for the spheroids.
- Invert the lid of the 10 cm3 dish to allow the media, including the cell suspension, to hang over a humid environment. Leave the hanging droplets for 3 days.
- Take images of the spheroids using an inverted microscope from day 3 until day 10 of culture.
- Put the 10 cm3 dish on the microscope stage. Adjust the magnification of the microscope at 50x for all spheroids.
- Open the microscope software on the attached computer and adjust its focus to have a clear image of every spheroid. Use the Capture Tool on the microscope software to save the acquired pictures.
7. Homotypic Hep3B spheroids in LX2 CM
- Suspend 3000 Hep3B HCC cells in the hanging droplets to form spheres. Add 10 mL of sterile PBS to the bottom of the dish to provide humid conditions for the spheroids.
- Invert the lid of the 10 cm3 dish to allow the media, including the cell suspension, to hang over a humid environment. Leave the hanging droplets for 3 days.
- Transfer Hep3B spheroids into 20 µL of fresh CM from LX2 cells in hanging droplets.
NOTE: Use sterile autoclaved unfiltered 200 µL pipette tips in the process of spheroid transfer to avoid any disruption or injury to the formed spheroids. This is also to eliminate any residual cells that remain unattached to the main single spheroid.- Use an autoclaved 20 µL pipette for the transfer process. Adjust the pipette volume to 2 µL. Attach the pipette tip to the pipette.
- Invert the lid of the 10 cm3 dish on which the spheroids were formed. Fix the lid on the stage of a light microscope. Adjust the fine focus of the microscope to make each spheroid visible.
- Carefully empty the air from the micropipette by pressing the plunger button. Insert the pipette tip in the droplet, including the spheroid, to be transferred. Get very close to the spheroid without touching it with the tip.
- Gently release the pressure on the plunger button to allow the suction of the spheroid into the micropipette tip in 2 µL media.
- Transfer the spheroid into a new droplet hanging on a new 10 cm3 dish, having new media/conditioned media/treatment.
NOTE: Ensure that all the spheroids are successfully transferred to the new 10 cm3 dish using the light microscope.
- Take images of the spheroids at 50x magnification using an inverted microscope from the day of transfer (day 3) until day 7 of culture in LX2 CM.
8. Calculation of spheroid volume
- Assign a unique numeric identifier for each spheroid so that images from matched spheroids can be captured daily.
- Analyze images of the growing spheroids using an image analysis software package.
- Open each spheroid image within the software package. Using the Freehand selection tool and outline each spheroid. From the Analysis dropdown button, select Set Measurement, and then Area. Press OK.
- Manually draw a circle around each spheroid. Once the sphere is circled, press Ctrl + M to allow the program to calculate the spheroid area in Pixels. Convert the area of the spheroid into a volume.
NOTE: Volume of spheroidmm3 =0.09403 ×
- Calculate the change in spheroid volume relative to its volume on the first day of image capture.
NOTE: This is to normalize the spheroid volume to the starting volume and improve accuracy given the natural variation in the starting size.
Cells cultured in a multi-layered 3D format more accurately reflect the complexity of the tumor microenvironment than conventional 2D cultures24,25. Previously, many studies have supplemented the spheroids culture media with different mitogens and growth factors26 to initiate spheroid formation. In this study, however, the addition of fibroblasts, or their CM, provides essential mitogens and growth factors to accelerate spheroid growth.
Figure 1 depicts data from a pilot study in which Huh7 human HCC cells were seeded in a descending seeding density starting from 12,000 down to 125 cells in 20 µL fresh media for 3 days. A seeding density of 12,000 cells yielded spheroids with an asymmetric shape, which was not corrected even after halving the cell seeding density. However, spheroids created from 3000 cells appeared more rounded (Figure 1, upper row). Further reduction of cell concentration neither succeeded in forming single spheres (125, 250, 500 cell-density spheres) nor had a regular spherical-like appearance (1000 and 1500 cell-density spheroids). It is worth mentioning that many small spheres were formed at a density of 500 tumor cells; however, only one small spheroid was captured at 50x magnification (Figure 1, upper row). The same optimized protocol was applied to the COS7 primate kidney fibroblast cell line (Figure 1, middle row). Suspending 125, 250, and 500 COS7 fibroblasts in 20 µL hanging droplets resulted in one rounded spheroid with multiple smaller spheroids, while higher cell densities (1000, 1500, and 3000) formed single semi-rounded spheroids. Similar to the Huh7 tumor spheroids, higher cell suspensions (6,000 and 12,000 cells/sphere) resulted in the formation of irregular cell aggregates, and hence higher cell concentrations were discarded (Figure 1, middle row). Low densities of Huh7/COS7 heterotypic cell suspensions (125, 250, and 500 cells/sphere) generated a single spheroid with multiple floating or semi-attached spheroids (Figure 1, bottom row). 1000- and 1500-cell heterotypic spheroids were semi-rounded, while a seeding density of 3000 cells (1500 per cell type) gave a rounded 3D spheroid. As noted earlier, higher cell densities resulted in the formation of aggregates rather than well-defined spheroids (Figure 1, bottom row). In conclusion, 3000 cell-density spheroids were rounded, similar to human HCC tumors, and were adapted for further experiments. Culturing the cell suspensions as hanging droplets for 3 days was sufficient to promote spheroid formation.
After optimizing the best cell concentration and time point for spheroid formation in the current context, the proliferative impact of co-culturing tumor and fibroblast cell lines was assessed longitudinally. Figures 2 and Figure 3 show Huh7/COS7 heterotypic spheroids (3000 total cell numbers per spheroid) monitored from day 3 until day 10. Homotypic Huh7 and COS7 spheroids (1500 cells each) served as controls. Starting from day 4, heterotypic spheroids grew in an ideal round-like shape compared to homotypic spheroids (Figure 2). The initial volume of the heterotypic spheroid was larger than that of each homotypic spheroid. The growth of spheroids was calculated as the change in their volume relative to initial volume at the day of spheroid formation to avoid normal variation in spheroid volume (day 3, Figure 3). Heterotypic spheroids initially showed a rapid growth phase starting from day 4 up to day 7 followed by a slower phase of growth at day 8 (Figures 2, upper row, and Figure 3). The spheroid volume decreased on days 9 and 10, possibly reflecting the depletion of nutrients or a hypoxic core and cell death.
In contrast, homotypic Huh7 and COS7 spheroids grew at a much slower rate (Figure 2, middle and bottom rows; Figure 3). Homotypic spheroids exhibited a relatively static growth curve until the fifth day of culture (two days after spheroid formation). Starting from day 6, the homotypic spheroids began to show a gradual increase in their growth curve, albeit at a significantly lower rate than that of the heterotypic spheroids (Figure 3). In conclusion, tumor/fibroblast heterotypic spheroids grow at a higher rate than homotypic spheroids suggesting that the direct contact of tumor cells and fibroblasts increases the size of the tumor spheroids.
Finally, to validate the above findings and to study the paracrine impact of mesenchymal cells on the proliferation of HCC spheroids in a liver-related context, 3-day-old homotypic Hep3B HCC spheroids were grown in regular fresh media or CM from LX2 hepatic stellate cells (Figure 4 and Figure 5) for an additional 4 days. At first glance, 3000 Hep3B cells formed perfectly rounded spheroids after 3 days (Figure 4). Hep3B spheroids showed continual proliferation in fresh media from day 3 till day 7 (Figure 4, upper row). The growth rate was enhanced when Hep3B spheroids were maintained in LX2 CM (Figure 4, lower row). This media-dependent conversion in Hep3B spheroid growth exhibited statistical significance from day 4 up to the end of the experiment (Figure 5), suggesting a fibroblast-driven proliferation of tumor spheroids.
In conclusion, this study has successfully modified the existing 3D spheroid cultures to exploit the crosstalk between HCC tumor cells and different fibroblast cell lines and investigate the proliferative significance of this direct and indirect cellular interaction (Figure 6). Further characterization of the formed spheroids is needed to improve how tumor cells interact with the surrounding microenvironment.
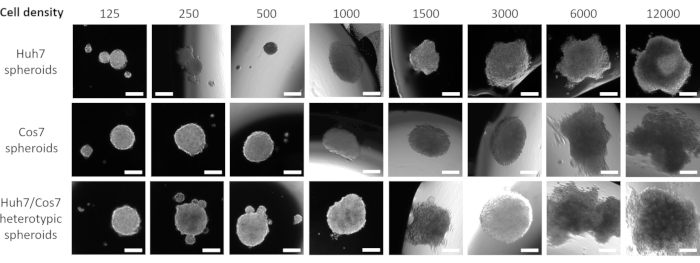
Figure 1: Optimization of the optimal cell density for spheroid formation. Columns represent different cell seeding densities, and rows represent spheroids formed from Huh7 cells (top row), COS7 cells (middle row), and Huh7/COS7 heterotypic cells (bottom row). Images represent spheroids formed after suspending 125, 250, 500, 1000, 1500, 3000, 6000, and 12000 cells (from left to right) in 20 µL media for 3 days. The pilot study included two spheroids per condition, and the images were taken at 50x magnification, scale bar = 200 µm. Please click here to view a larger version of this figure.
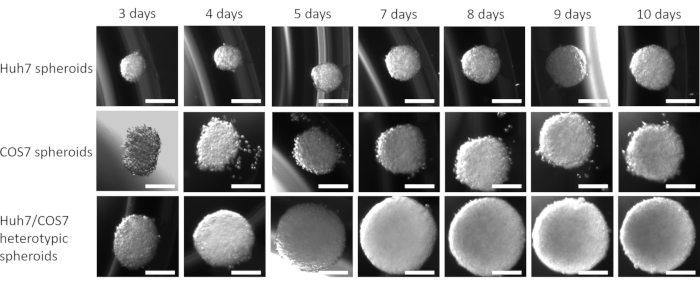
Figure 2: Heterotypic versus homotypic spheroid growth. Columns represent the day at which spheroid images were taken, and rows show representative images for spheroids formed from Huh7 cells (top row), COS7 cells (middle row), and Huh7/COS7 cells (bottom row). The images represent the spheroids formed after suspending 3000 cells from each condition (homotypic Huh7, COS7, or the Huh7/COS7 heterotypic spheroids) of different time points from left to right. The experiment included 10 spheroids per condition, and the images were taken at 50x magnification, scale bar = 200 µm. Please click here to view a larger version of this figure.
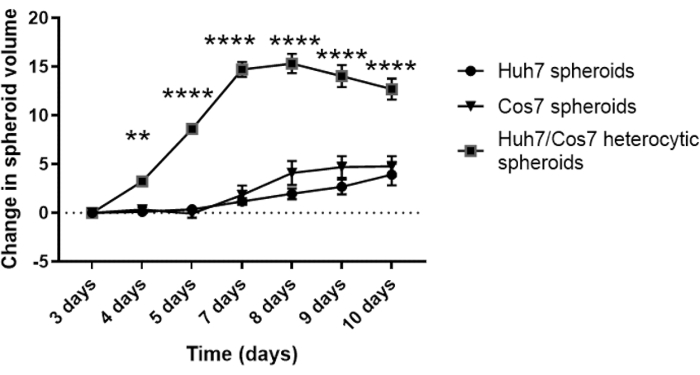
Figure 3: Longitudinal analysis of heterotypic versus homotypic spheroid growth. The graph shows the growth curve of heterotypic Huh7/COS7 spheroids versus homotypic Huh7 and COS7 spheroids. Data are presented as mean ± s.e.m; n = 10 independent spheroids. ** p < 0.01; ****p < 0.001. Please click here to view a larger version of this figure.
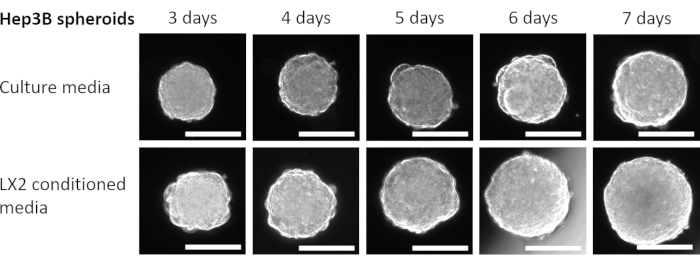
Figure 4: Growth of Homotypic Hep3B spheroids in LX2 CM. Columns represent the day at which spheroid images were captured, and rows show representative images of Hep3B spheroids cultured in fresh DMEM culture media (top row) or LX2 CM (bottom row). The experiment included seven spheroids per condition (fresh media or LX2 CM), and images were taken at 50x magnification, scale bar = 200 µm. Please click here to view a larger version of this figure.
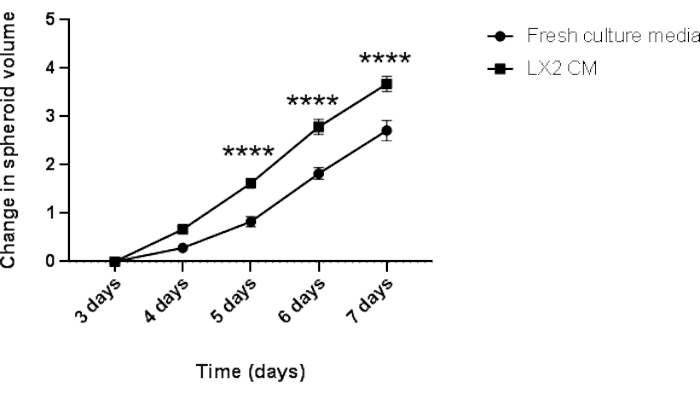
Figure 5: Longitudinal analysis of the homotypic Hep3B spheroid growth. The graph shows the growth curve of Hep3B spheroids in fresh DMEM culture media or LX2 CM. Data are presented as mean ± s.e.m; n = 7 independent spheroids. ****p < 0.001. Please click here to view a larger version of this figure.
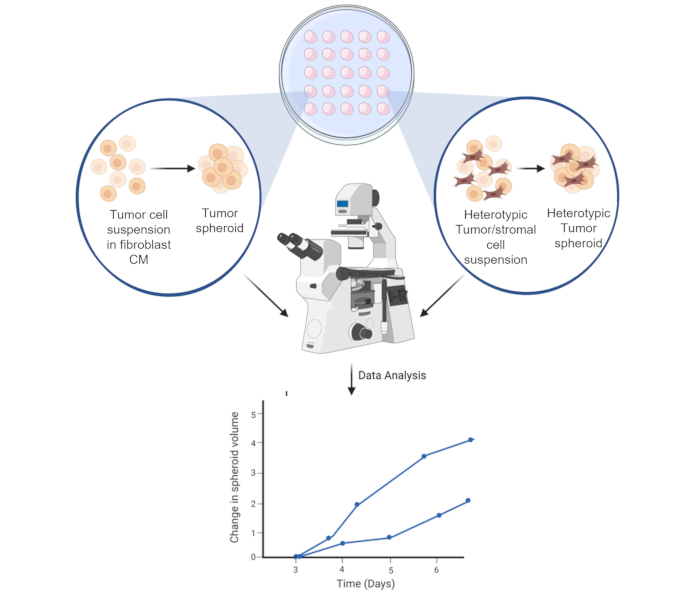
Figure 6: Schematic representation of the spheroid formation process. The cell suspension is pipetted on the inner lid of 10 cm3 Petri-dish. The lid is inverted and kept for 3 days to allow for homotypic or heterotypic spheroid formation. Spheroid images are taken at 50x magnification. The figure is created using a web-based science illustration tool (see Table of Materials). Please click here to view a larger version of this figure.
