The media used to grow bacterial biofilms influence the survival of neutrophils. Different media were tested to reduce the effect of media alone on the viability of neutrophils for studying neutrophil-biofilm interactions (Figure 1). Bacterial growth media such as Tryptic Soy Broth minimizes the viability of neutrophils, such that ~60% of neutrophils are alive after a 30 min incubation period at 37 °C with 5% CO2. Mammalian cell culture media, such as MEMα, does not affect the viability of neutrophils and supports the growth of S. aureus biofilms. In fact, minimal media promotes robust growth of biofilms in other bacteria46,47.
To assess the effect of media on biofilm growth and variability in biofilm biomass quantification after washing the biomass to eliminate planktonic cells, an 18 h S. aureus biofilm was grown in a 96-well plate, with wells treated or untreated with poly-L-Lysine. A nutrient-rich (Tryptic Soy Broth (TSB)) and minimal (MEMα) media were used as-is or supplemented with 2% glucose. The biofilm biomass stained with CV revealed that S. aureus biofilm grown in MEMα supplemented with 2% glucose produced the most robust biofilm among all tested media (Figure 2A). Furthermore, biofilms grown in PLL pretreated wells containing MEMα + 2% glucose showed less variability than biofilms in PLL-untreated wells containing MEMα + 2% glucose. These biofilms showed less variability in quantification via CV assay35 and the CFU/mL when plated after precisely handling biofilms for biomass quantification. These biofilms contained, on average, 1 x 108 CFU/mL, as demonstrated by plating the biofilms in 3 separate days (Figure 2B). This number is useful in determining the number of neutrophils to add to the biofilms for neutrophil functionality assays.
To measure ROS production by neutrophils in response to biofilms, S. aureus biofilms were grown statically for 18-20 h in a 96-well plate. Biofilms were then opsonized, and neutrophils were added. ROS production was then measured for 60 min (Figure 3A). The area under the curve is calculated from the kinetic curve to quantify total ROS production by neutrophils. Neutrophils treated with an agonist, such as PMA, used as a control, show an increased ROS production. In the absence of biofilms, neutrophils treated with PMA showed robust ROS production. In the presence of S. aureus biofilm, the overall ROS production by neutrophils treated with PMA decreased. In the absence of PMA, neutrophils solely rely on their interaction with the biofilm, which further reduces the amount of ROS produced (Figure 3B).
To visualize the neutrophil-biofilm interactions using fluorescence microscopy, a GFP-expressing strain of S. aureus, Blue CMAC dye, and ethidium homodimer-1, that stains the cytoplasm of live cells and DNA of dead cells, respectively, were used. S. aureus biofilm was grown for 18 h in a 6 µ-channel slide. Blue CMAC dye-labeled neutrophils were added along with ethidium homodimer-1 to the washed biofilms and incubated for 30 min at 37 °C with 5% CO2 prior to imaging. Wide-field fluorescent microscopy revealed that many neutrophils were localized to the surface of S. aureus biofilms, while a few are within the biofilm (Figure 4A). The interaction between S. aureus cells within neutrophils was also apparent (Figure 4C). Most of the S. aureus cells interacting with neutrophils (cyan) were dead (magenta), while a few remained alive (yellow) as determined by live-dead staining (Figure 4C). For comparison, GFP-expressing S. aureus biofilms were stained with ethidium homodimer-1, which revealed a fraction of the dead S. aureus population within the biofilm (Figure 4B). Non-viable neutrophils that were positive for ethidium homodimer-1 were quantified using analysis software (see Table of Materials) after incubation with S. aureus biofilms. Approximately 48% of neutrophils were already dead within 30 min of incubation with S. aureus biofilm. During optimization of the microscopy protocol, the effect of washing the biofilm and neutrophils after 30 min of incubation to remove non-adhered neutrophils was also assessed, revealing around 33% of dead neutrophils still attached to the biofilm (Figure 4D).
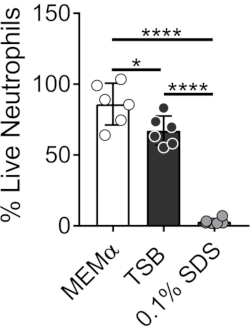
Figure 1: LIVE-DEAD assay compares neutrophil survival between bacterial and mammalian growth media. Neutrophils were isolated and incubated in HBSS, MEMα, TSB, or 0.1% SDS for 30 min. LIVE-DEAD staining was performed using Calcein AM (live) and ethidium homodimer-1 (dead). Percent of live neutrophils was determined, where HBSS-incubated neutrophils were treated as 100% live neutrophils. Results represent an average of two independent experiments performed in triplicate, with neutrophils obtained from two different donors. Data are presented as mean ± SD (*p < 0.05, ****p < 0.0001. One-way ANOVA). Please click here to view a larger version of this figure.
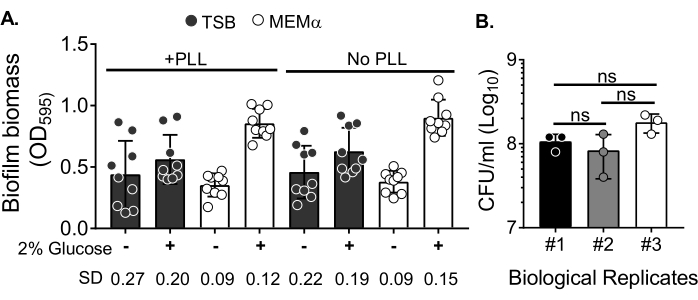
Figure 2: Quantification of biofilm biomass in different conditions and bacterial viability count of biofilms grown in the optimized conditions. (A) S. aureus was seeded in a 96-well plate either coated or uncoated with poly-L-Lysine (PLL). Biofilms were grown in TSB, MEMα, or either of the media supplemented with 2% glucose under static conditions for 18 h. Crystal violet (CV) staining was performed to stain biofilm biomass. The eluted CV stain was diluted at 1:10 and read in a microplate reader. Results represent an average of three independent experiments performed in triplicate. Data are presented as mean ± SD. The SD for each group is shown at the bottom to demonstrate different biofilm growth conditions variability. (B) Bacterial CFU counts were obtained from biofilms grown in an optimized medium (MEMα + 2% glucose). The 18 h static biofilms were subjected to the same number of washes followed by a 10 min sonication to loosen the biofilm biomass and passed through a 22G needle to disrupt the aggregates prior to plating. Results represent three replicates performed in triplicate. Data are presented as mean ± SD (ns = not significant. One-way ANOVA). Please click here to view a larger version of this figure.
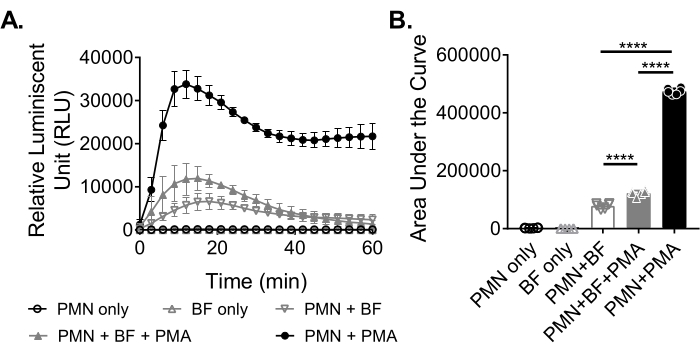
Figure 3: Quantification of ROS production by neutrophils via chemiluminescence assay. (A) Neutrophils (PMN) were incubated with HBSS-washed S. aureus biofilms (BF) either in the presence (closed gray triangle) or absence (open gray inverted triangle) of PMA to measure ROS production by neutrophils. Luminol was used to detect ROS every 3 min for 60 min in a microplate reader. While neutrophils treated with PMA in the absence of a biofilm (closed black circle) served as a positive control, neutrophil only (open black circle) and biofilm only (open gray triangle) groups served as negative controls. Data represent an average of two independent experiments performed in triplicate with neutrophils obtained from two different donors. Data are presented as mean ± SD. (B) The area under the curve from (A) was calculated to quantify the total ROS generated by the neutrophils. The data are represented as mean ± SD. (***p < 0.0001. One-way ANOVA). Please click here to view a larger version of this figure.
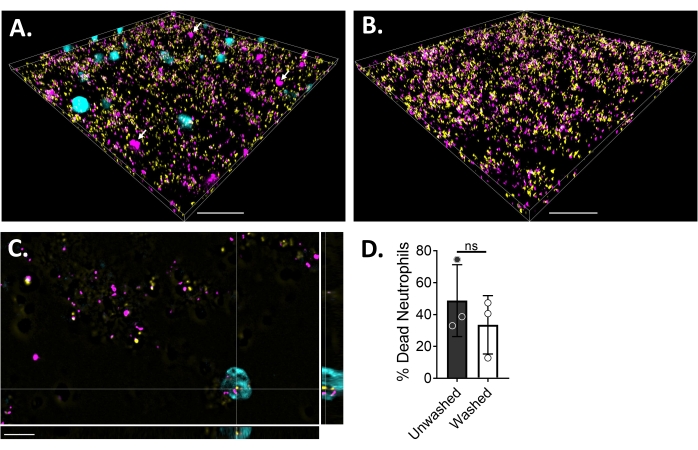
Figure 4: Visualization of the interaction between S. aureus biofilm and neutrophils using wide-field fluorescence microscopy. Blue CMAC dye-labeled neutrophils (cyan) were supplemented with ethidium homodimer-1 (magenta; dead) prior to incubating with an 18 h S. aureus biofilm (yellow). Biofilm-neutrophil interactions were imaged using wide-field fluorescent microscopy and images processed using an image analysis software. Experiments were performed with three different donors. Representative images are presented as (A) 3D view of S. aureus biofilm with live (cyan) and dead (magenta; a few indicated with white arrows) neutrophils, (B) 3D view of an S. aureus biofilm in the absence of neutrophils with either live S. aureus expressing GFP (yellow) or dead S. aureus stained with ethidium homodimer-1 (magenta), (C) an orthogonal view of S. aureus and neutrophil interaction as depicted by the xy, yz, and xz planes, and (D) quantification of neutrophil viability in the presence of S. aureus biofilm after 30 min either immediately (unwashed) or after three rounds of washes with HBSS to remove non-adhered neutrophils (washed). Neutrophil cell death is presented as mean ± SD (Student's t-test). Scale bar indicates 50 µm in (A) and (B) and 10 µm in (C). Please click here to view a larger version of this figure.
