In this work, WFISH was used to investigate the chromosome behavior during spermatogenesis in An. gambiae. The first crucial step for applying this protocol is obtaining testes that show a low level of morphological alteration after dissection. Basic knowledge of the mosquito anatomy is required to perform a successful testis dissection, and below, some guidances for this procedure are given. In the Anopheles mosquito, mature testes can be found lying in the sixth abdominal segment of the pupal and adult stages21. As shown in Figure 1, the vas deferens connects the testes to the male accessory glands (MAGs), which are located in the last segment of the abdomen. The MAGs are connected to a unique ejaculatory duct that delivers the sperms and seminal fluids to the copulatory organ and the external part of the male genital apparatus21. The entire internal male genital apparatus can be dissected using different approaches depending on the life stage of the mosquito. During the pupal stage, testes can be easily identified using a stereomicroscope throughout the light cuticle by looking at the ventral side of the abdomen in the proximity of the sixth segment (Figure 1). To dissect the testes, the lower part of the abdomen, including the sixth segment, can be isolated from the rest of the body using a pair of needles and transferred into a clean drop of 1x PBS. Following the removal of the last segment, the entire apparatus can be squished out of the abdomen by applying gentle pressure with the dissecting needles. To dissect the testes from adult males, the first step involves isolating the entire abdomen in a fresh drop of 1x PBS and then pulling out the last segment carrying the clasps, which are the male copulatory structures (Figure 1). At this point, the lower part of the MAGs should emerge and be easily identifiable due to their yellow color. The entire male apparatus can then be slowly pulled out with the help of a needle or forceps in a drop of 1x PBS until the pair of testes attached to the vas deferens is visible. Before proceeding with the fixation, it is important to isolate the testes from the other parts of the male apparatus by cutting in the proximity of the lower part of the vas deferens (Figure 1).
The age of the pupae or adult males is an important factor to consider depending on the spermatogenesis stage under investigation. In An. gambiae, spermatogenesis starts at the early/mid-pupal stage, and it continues throughout the entire life of the individual24. Between 3 h to 10 h after pupation, the premeiotic and meiotic stages are more represented in the testis (meiotic prophase, meiotic divisions), the spermatid DNA is relatively uncondensed, and mature sperms have not yet been formed. Late pupae and 1 day old adults offer a good balance between the premeiotic, meiotic, and post-meiotic stages (Figure 2 and Figure 3). In adults that are more than 4 days old, the premeiotic stages and spermatocysts are less represented, and the testes are mainly occupied by mature sperms contained in the sperm reservoir.
To investigate the behavior of the sex chromosomes during different stages of spermatogenesis, WFISH was performed on testes dissected at the late pupal stage to ensure a good representation of the entire process. To follow the behavior of these chromosomes, fluorescent probes specific to repetitive sequences located exclusively on the X or Y chromosome were used. The fluorescent probes can be generated using PCR or obtained commercially as 3' end-labeled oligonucleotides. Using an oligo with a length of >40 bps is recommended to allow good signal detection from the fluorescent oligo probe. In our experience, 3' end-labeled oligos perform better than PCR-labeled probes in terms of signal detection. In addition, the copy number of the target sequence is a factor that may affect the efficacy of WFISH. If labeling is unsuccessful, using the PCR labeling method on a longer fragment or designing several oligos specific to the target region is suggested.
The current methods, based on using a penetrating solution (1% Triton/0.1 M HCl in 1x PBST), allow a good level of testis permeabilization and penetration of the probes, resulting in a successful hybridization reaction. Oligo probes specific to sex chromosome repetitive sequences can be designed based on the extensive characterization of repetitive elements performed by Hall et al.20. In addition, consensus sequences specific to X- or Y-linked repetitive elements can be obtained using a bioinformatic platform such as the RedKmer pipeline30. It is important to notice that sex chromosome probes can target repetitive elements such as satellites and retrotransposons, and they can have a different level of hybridization with the X or the Y chromosomes depending on the species under examination20,31,32. As shown in Figure 3, a good level of hybridization of the probes and low background allowed the visualization of the targeted chromosomes throughout spermatogenesis. The pairing of labeled sex chromosomes could be seen in the premeiotic and meiotic stages. This was followed by detecting either the X or the Y chromosomes in haploid cell nuclei chromosomes resulting from meiotic divisions. Subsequently, X- or Y-bearing spermatids could be followed throughout spermiogenesis, marked by different levels of DNA condensation, to the final step of arrow-shaped mature spermatozoa. In the present experimental setup, confocal Z-stacks were used to acquire information regarding the 3D spatial organization of the cells during this process (Video 1).
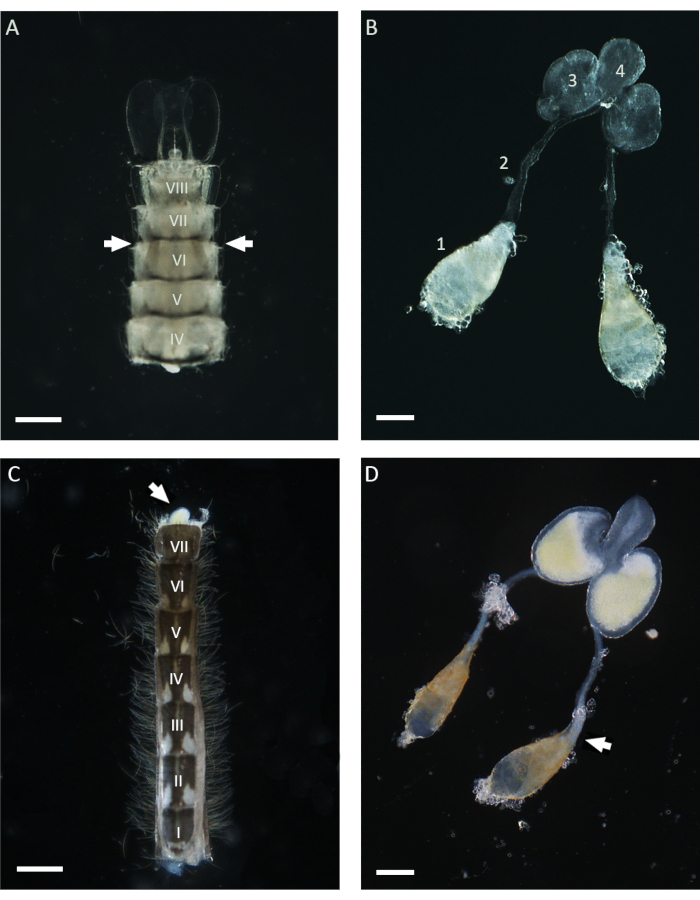
Figure 1: Testes dissected from pupae and 1 day old adult Anopheles gambiae males. (A) An abdomen dissected at the late pupal stage showing the position of the testes in the proximity of the sixth abdominal segment. The testes can be identified throughout the cuticle and appear as brownish structures on both sides of the abdomen (with arrows). (B) Testes dissected at the pupal stage, showing the mature testes (1), vas deferens (2), MAGs (3), and ejaculatory duct (4). (C) An abdomen dissected from a 1 day old adult male after removing the basal clasp segment. The MAGs can be squished from the abdomen by applying gentle pressure (white arrow). (D) Male internal reproductive apparatus dissected from a 1 day old adult male. The white arrow indicates the position occupied by mature sperms, which appear as a white aggregate at the basal pole of the testis. Scale bars: (A,C) 200 µm; (B,D) 100 µm. The Roman numerals from I to VIII indicate the abdominal segments. Please click here to view a larger version of this figure.
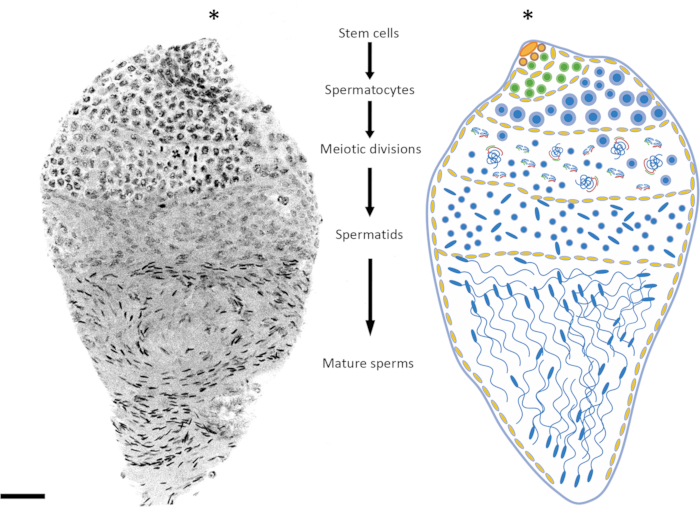
Figure 2: Representation of spermatogenesis in Anopheles gambiae. The image on the left shows an An. gambiae late pupa testis following whole-mount DAPI staining. On the right is a schematic version for better visualization. By observing the nuclear shape and condensation level, it is relatively easy to follow all the spermatogenesis stages from diploid cells to haploid spermatozoa. The stem cell niche is situated in the upper pole of the organ, where differentiation into spermatogonia starts. The spermatogonia cells increase in number after mitotic division (green cells), and the spermatocysts increase in size (yellow cells). The spermatogonia cells differentiate into spermatocytes after multiple rounds of mitotic divisions (blue cells). The spermatocytes, which are characterized by relatively larger nuclei than the cells of the other stages of the process, are the cells that will undergo meiotic division. Cells undergoing meiosis can be detected by looking at the presence of chromosomes at different meiotic stages; chiasmata and metaphase chromosomes can be detected even at low magnification. The premeiotic stages are overrepresented in testes dissected at the early pupal stage. After the first and second meiotic divisions, spermatids are produced and can usually be found lying in the middle of the testis. The nuclei of spermatids show a certain degree of variation in their shape, from a round to an arrow-like shape. The spermatids enter the spermiogenesis process, during which the nuclei start condensing, and their structure changes into arrow-like dots. When mosquitoes mature sexually after emerging, spermatocysts containing mature sperms can occupy most of the testis volume at the expense of spermatocysts at a different stage of development. Scale bar: 20 µm. The asterisk (*) indicates the apical part of the testis. Please click here to view a larger version of this figure.
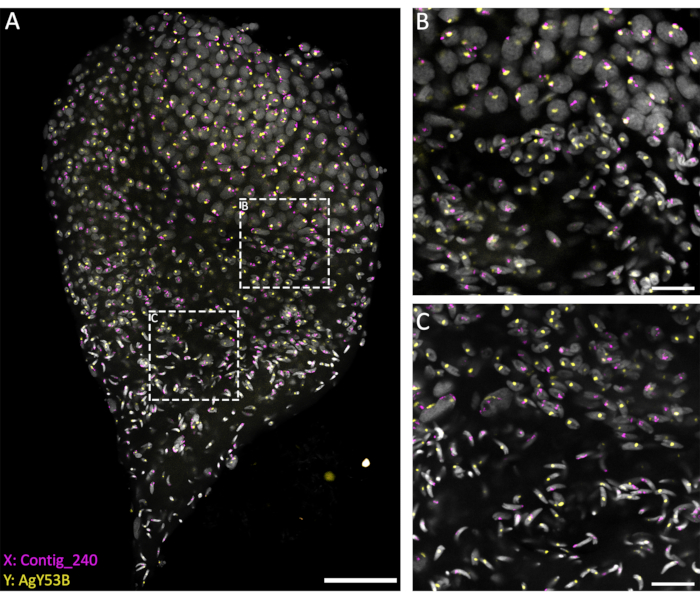
Figure 3: WFISH on an An. gambiae testis dissected from the late pupal stage. WFISH was performed using probes specific for the X (Contig_240) and Y (oligo probe specific for AgY53B) chromosomes. (A) WFISH allows to follow the behavior of the sex chromosome during spermatogenesis from diploid cells to haploid spermatozoa. In this image, it is possible to appreciate the dramatic changes that the nuclei undergo during spermatogenesis. Labeling the sex chromosomes allows for the discrimination between diploid and haploid cells. In diploid cells, the signal from the sex chromosomes is linked to the same nuclei. In haploid cells (spermatids and spermatozoa), the signal of the sex chromosomes is unlinked due to the meiotic reductional division. (B,C) A higher-magnification (63x) image of the testis shown in (A). They were acquired at different positions along the Z-axis. The white dotted frames indicate the area of acquisition. (B) The transition stage between spermatocytes and spermatids, showing the formation of haploid cells and the separation of the signals from the sex chromosomes into separate nuclei. (C) The transition stage between haploid spermatids and mature spermatozoa. This stage shows the changes in the nuclear condensation level; mature spermatozoa show a more condensed and elongated shape than spermatids. Scale bars: (A), 30 µm; (B,C), 10 µm; Gray: DAPI. Please click here to view a larger version of this figure.
| Target sequence | Primer Sequence and Oligo-probe consensus | Reference |
| Contig_240 (X) | 5’-CAATAAATTTCCTTTTTAATGATGC AAAATCTACGTCTCTAGC-3’-[Fluorochrome] |
19 |
| AgY53B (Y) | 5’AGAAGAATAGAATCAGAATAGTCGG TTTCTTCATCCTGAAAGCC-3’-[Fluorochrome] |
This study |
| AgY477- AgY53B junction region (Y) |
5’-TTCTAAGTTTCTAGGCTTTAAGGAT GAAGAAACCGACTATTC-3’-[Fluorochrome] |
19 |
| 18S rDNA (X) | F: AACTGTGGAAAAGCCAGAGC R: TCCACTTGATCCTTGCAAAA |
19 |
| AgY53B (Y) | F: CCTTTAAACACATGCTCAAATT R: GTTTCTTCATCCTTAAAGCCTAG |
19 |
Table 1: List of oligo probes specific to the X or Y chromosome in An. gambiae.
Video 1: A 3D stack on WFISH performed on an An. gambiae testis dissected from the late pupal stage. To obtain a 3D representation of the spermatogenesis process, a confocal 3D stack can be performed on testes showing a low number of structural alterations. In this study, the stacks were performed with an interval of 1.25 µm between two optical sections under a 63x or 40x oil lens in order not to lose information about the 3D spatial organization of the cells. Gray: DAPI, yellow: Contig_240 (X), magenta: AgY53B (Y). Please click here to download this Video.
