The apical region of the adult mouse incisor is encased within the mandible (Figure 1) and hence, not directly accessible for visualizing and live-tracking the progenitor cells residing within the growth region. Therefore, we have developed a method to extract the whole incisor from the jawbone and maintain it in an explant culture system for two-photon timelapse microscopy (Figure 2). Here we describe representative results that capture the dynamic process of cell proliferation and movement in the labial cervical loop region of the dental epithelium.
To demonstrate the experimental procedures, we have used two different mouse models that express green fluorescence in the dental epithelium. The first mouse line is K14Cre;R26rtTA;tetO-H2B-GFP, where K14Cre (MGI:2680713) expresses the Cre recombinase from a Keratin 14 promoter35 and activates the expression of the reverse tetracycline-controlled transactivator in the epithelium from the R26rtTA allele36 (MGI:3584524). Upon administration of doxycycline, rtTA induces histone H2B-GFP expression from the tetO-H2B-GFP allele37 (MGI:3043783) and labels epithelial cell nuclei with green fluorescence. This is especially useful for cell tracking and for detecting cell divisions. In this experiment, we fed animals with doxycycline food for 24 h before sacrifice to activate H2B-GFP expression. The second mouse line is K14Cre;R26mT/mG, in which R26mT/mG (MGI:3803814) is a Cre-reporter38. In the absence of Cre activity, cells express membrane-localized tdTomato (mT) with red fluorescence. Upon Cre-mediated recombination, cells express membrane GFP (mG). K14Cre;R26mT/mG thus labels epithelial cell membranes with green fluorescence, leaving non-epithelial cells red. This permits easy visualization of cell shapes, divisions, and movements.
We began the procedure by dissecting out the mandibles (Figure 3A) and then systematically removed all the bones surrounding the incisor (Figure 3B–G). This yielded whole incisors with undamaged epithelium (Figure 3H). We confirmed the intactness of the dental epithelium by inspecting the green fluorescence in the K14Cre;R26rtTA;tetO-H2B-GFP and K14Cre;R26mT/mG mice (Figure 4A,B,E,F). At this stage, the opaque periodontal tissues still cover the apical incisor, and the cervical loop thus appears blurry due to light scattering, which would similarly hinder downstream timelapse imaging (Figure 4C,G). We therefore carefully removed the periodontal tissues, so the dental epithelium with the cervical loop could be discerned in each incisor (Figure 4D,H).
The incisors were then embedded in low melting point agarose and cultured in a perfusion setup for two-photon live imaging as depicted in Figure 2. For this exercise, we have focused on the cervical loop region of the dental epithelium and captured z-stack images at 4 µm intervals every 5 min over a duration of 14 h (Figure 5A). Notably, H2B-GFP signals were mostly observed in the transit-amplifying region of the cervical loop, where there are active cell divisions (Figure 5B). This is likely because there is higher H2B-GFP exchange at open chromatins and incorporation into nucleosomes following DNA replications in these active cells39.
We subsequently examined the timelapse images using ImageJ and based on the separating of the H2B-GFP signals40, we were able to observe numerous cell divisions throughout the imaging period (Supplemental Video S1 and Supplemental Video S2). This indicated that the tissues were adequately maintained in the explant culture and cells were active. Specifically, we were able to observe the condensation and alignment of chromosomes at the metaphase plate in mitotic cells, followed by their segregation into two daughter cells during anaphase (Figure 5C–N, manually tracked using the ImageJ plugin TrackMate). Most of these divisions were perpendicular or at an oblique angle relative to the basement membrane (Figure 5C–K and Supplemental Video S1). Horizontal divisions parallel to the basement membrane could also be detected, although occurring less frequently (Figure 5L–N and Supplemental Video S2). It is important to point out that cytokinesis in the dental epithelium often happens quickly, within 5-10 min. As a result, timelapse intervals of more than 5 min may miss some of these divisions.
Cell division events were also apparent in K14Cre;R26mT/mG cervical loops, where all epithelial cell membranes were labeled green (Figure 6A). We could identify mitotic cells by their cell rounding and then cytokinesis (Supplemental Video S3), and both vertical and horizontal cell divisions were observable (Figure 6B–G), thus similar to the results obtained using H2B-GFP. Together, these results demonstrate that this protocol can serve as a powerful tool to investigate cell behaviors in the incisor explants when combined with mouse genetic models that fluorescently label distinct subcellular structures.
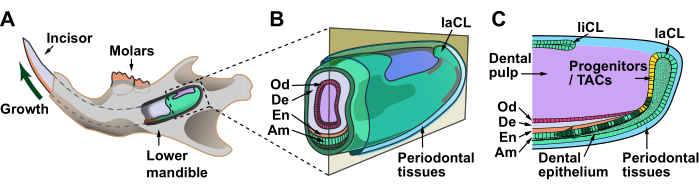
Figure 1: Schematics of the mouse jaw and incisor cervical loop. (A) A significant portion of the incisor is embedded in the jawbone. The growth region is located at the apical end of the tooth and supports its continuous growth (dark green arrow). (B) An enlargement of the apical incisor, which is surrounded by periodontal tissues. The tooth is composed of enamel and dentin, which are highly mineralized structures formed by ameloblasts and odontoblasts, respectively. (C) A sagittal section of the apical incisor, showing that dental epithelial progenitor cells and transit-amplifying cells reside in the labial cervical loop and give rise to ameloblasts in the more distal epithelium (dark green dashed arrow). Compared to the labial cervical loop, the lingual cervical loop is smaller in size and does not normally form ameloblasts. Dental mesenchymal stem cells are present in the dental pulp (purple region) and give rise to odontoblasts. Abbreviations: En = enamel; De = dentin; Am = ameloblast; Od = odontoblast; TACs = transit-amplifying cells; laCL = labial cervical loop; liCL = lingual cervical loop. Please click here to view a larger version of this figure.
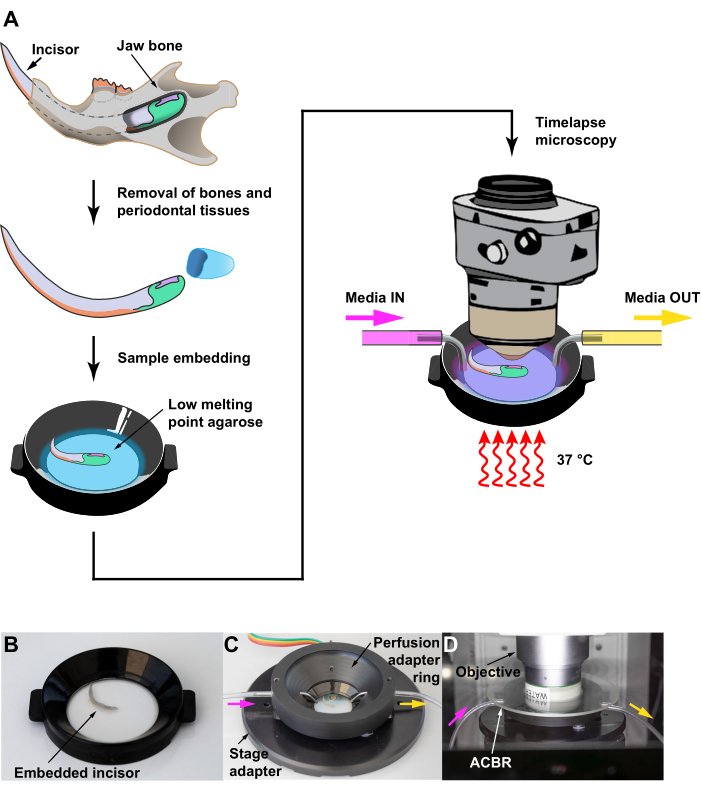
Figure 2: Maintaining mouse incisor explant for live imaging. (A) Schematics depicting key steps of the protocol, from dissecting the incisor to embedding the tissue in low melting point agarose for live imaging. A perfusion setup is used to provide a constant supply of nutrients during imaging and the culture is maintained at 37 °C. (B–D) A step-by-step demonstration of setting the culture dish and the perfusion chamber for live imaging. The media inlet and outlet are shown as pink and yellow arrows respectively. Abbreviation: ACBR = Atmospheric Control Barrier Ring. Please click here to view a larger version of this figure.
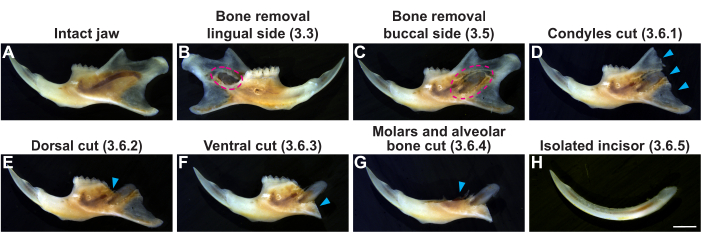
Figure 3: Isolation of the whole mouse incisor. (A) Intact jaw. (B–H) Bones were gradually removed to expose the whole incisor. Red dashed lines in B and C show the exposed soft tissue of the apical incisor after shaving off the membrane bone overlying the lingual and buccal sides of the incisor socket (steps 3.3-3.5 in the protocol). (D–G) Blue arrowheads represent the condyles, molars, and alveolar bones that have been removed to isolate the entire tooth (step 3.6 in the protocol). Scale bar = 2 mm (H). Please click here to view a larger version of this figure.
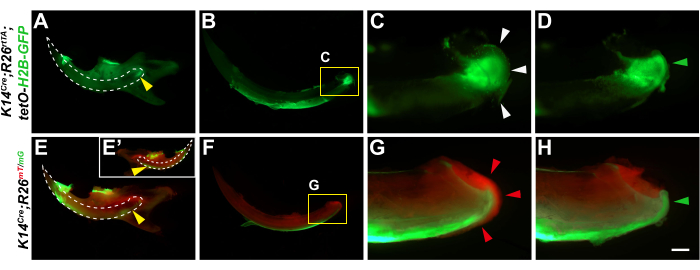
Figure 4: Removal of periodontal tissues for live imaging. (A–H) Representative samples from (A–D) K14Cre;R26rtTA;tetO-H2B-GFP,and (E–H) K14Cre;R26mT/mGmice were used in our demonstration of the protocol. Before dissection, GFP expression in the dental epithelium was visible through the bone (A,E, yellow arrowheads). White dashed lines outline the undissected incisors. E' shows the lingual side. In isolated incisors (B,C,F,G), GFP fluorescence was initially diffracted by the periodontal tissues covering the apical incisors (white and red arrowheads). Red fluorescence in F and G labels non-epithelial cells. Removal of periodontal tissues allows a clear and unobstructed view of the green fluorescent cervical loops (D,H, green arrowheads). Scale bar = 2 mm (A,E); 1.25 mm (B,F); and 300 µm (C,D,G,H). Please click here to view a larger version of this figure.
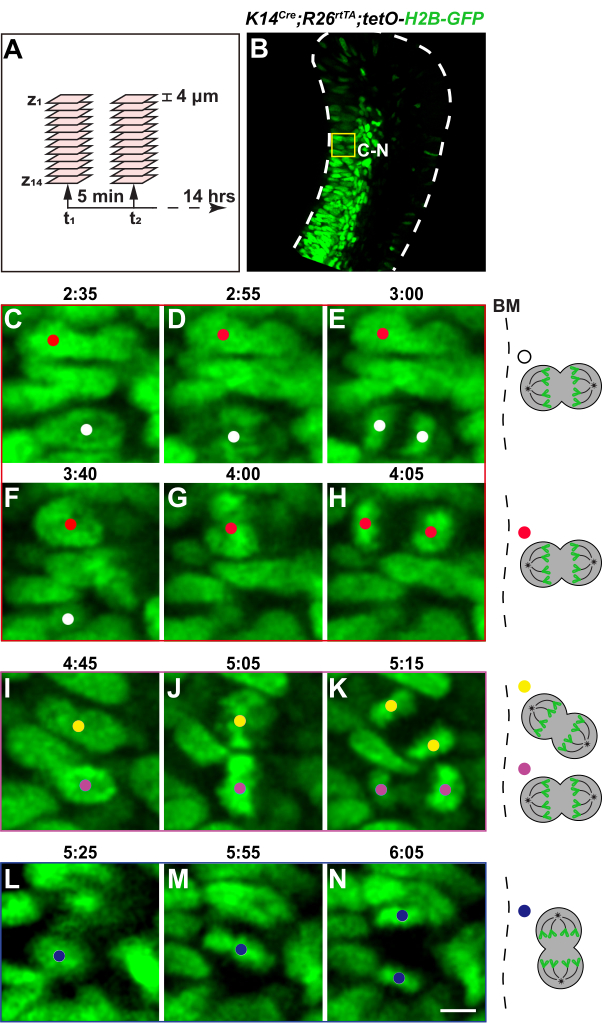
Figure 5: Timelapse microscopy of the K14Cre;R26rtTA;tetO-H2B-GFP cervical loop. (A) A schematic illustrating the timelapse setup. (B) A representative z-plane of the K14Cre;R26rtTA;tetO-H2B-GFP incisor labial cervical loop, showing nuclear labeling by H2B-GFP primarily in the transit-amplifying region, where cells actively divide. The yellow box represents the general area of the enlarged images shown below. (C–K) Timelapse images showing vertical and oblique cell divisions relative to the BM. (L–N) Timelapse images show an example of horizontal cell division relative to the BM. (D,G,J,M) Cells in metaphase or anaphase are displayed in the middle panels. Schematics of each tracked division are shown on the right. Scale bar in N = 36 µm (B); 5 µm (C–N). Abbreviation: BM = basement membrane. Please click here to view a larger version of this figure.
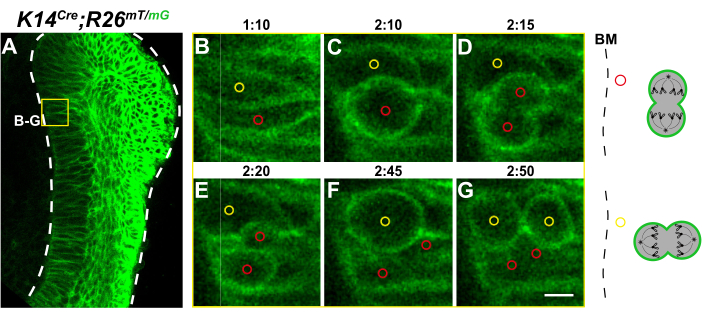
Figure 6: Timelapse microscopy of the K14Cre;R26mT/mG cervical loop. (A) A representative z-plane of the K14Cre;R26mT/mG incisor labial cervical loop. All epithelial cells express membrane GFP. The yellow box represents the area of cell tracking shown in the enlargements. (B–G) Timelapse images capturing both (B–D) horizontal and (E–G) vertical cell divisions, relative to the BM. (C,F) Middle panels show mitotic cell rounding. Schematics of each tracked division are shown on the right. Scale bar = 35 µm (A); 5 µm (B–G). Abbreviation: BM = basement membrane. Please click here to view a larger version of this figure.
Supplemental Figure S1: Inhouse-made perfusion dish. (A) A 35 mm culture dish can be used to make a perfusion dish. A glass bottom dish can be used if imaging is performed using an inverted microscope. (B) Heat a nail with a flame. (C) Two openings (white arrowheads) are created on either side of the dish using the heated nail. (D) Affix two blunt-end 16 G needles, one as the media inlet and the other as the media outlet, through the openings using epoxy glue. If gas perfusion is desired, a third opening/needle can be added to the dish. Please click here to download this File.
Supplemental Video S1: Live imaging of a K14Cre;R26rtTA;tetO-H2B-GFP incisor labial cervical loop, showing vertical and oblique cell divisions. Cells are manually tracked, and different colored dots represent different pairs of mother/daughter cells. Scale bar = 30 µm (left frame); 7 µm (right frame). Please click here to download this Video.
Supplemental Video S2: Live imaging of a K14Cre;R26rtTA;tetO-H2B-GFP incisor labial cervical loop, showing an example of horizontal cell division. The horizontal division takes place at the 10 s mark and is manually tracked using blue dots. Scale bar = 30 µm (left frame); 7 µm (right frame). Please click here to download this Video.
Supplemental Video S3: Live imaging of a K14Cre;R26mT/mG incisor labial cervical loop, showing both vertical and horizontal cell divisions. Cells are manually tracked, and different colored circles represent different pairs of mother/daughter cells. Scale bar = 30 µm (left frame); 7 µm (right frame). Please click here to download this Video.
