Direct Observation and Automated Measurement of Stomatal Responses to Pseudomonas syringae pv. tomato DC3000 in Arabidopsis thaliana
Summary
Here, we present a simple method for direct observation and automated measurement of stomatal responses to bacterial invasion in Arabidopsis thaliana. This method leverages a portable stomatal imaging device, together with an image analysis pipeline designed for leaf images captured by the device.
Abstract
Stomata are microscopic pores found in the plant leaf epidermis. Regulation of stomatal aperture is pivotal not only for balancing carbon dioxide uptake for photosynthesis and transpirational water loss but also for restricting bacterial invasion. While plants close stomata upon recognition of microbes, pathogenic bacteria, such as Pseudomonas syringae pv. tomato DC3000 (Pto), reopen the closed stomata to gain access into the leaf interior. In conventional assays for assessing stomatal responses to bacterial invasion, leaf epidermal peels, leaf discs, or detached leaves are floated on bacterial suspension, and then stomata are observed under a microscope followed by manual measurement of stomatal aperture. However, these assays are cumbersome and may not reflect stomatal responses to natural bacterial invasion in a leaf attached to the plant. Recently, a portable imaging device was developed that can observe stomata by pinching a leaf without detaching it from the plant, together with a deep learning-based image analysis pipeline designed to automatically measure stomatal aperture from leaf images captured by the device. Here, building on these technical advances, a new method to assess stomatal responses to bacterial invasion in Arabidopsis thaliana is introduced. This method consists of three simple steps: spray inoculation of Pto mimicking natural infection processes, direct observation of stomata on a leaf of the Pto-inoculated plant using the portable imaging device, and automated measurement of stomatal aperture by the image analysis pipeline. This method was successfully used to demonstrate stomatal closure and reopening during Pto invasion under conditions that closely mimic the natural plant-bacteria interaction.
Introduction
Stomata are microscopic pores surrounded by a pair of guard cells on the surface of leaves and other aerial parts of plants. Under ever-changing environments, regulation of the stomatal aperture is central for plants to control the carbon dioxide uptake required for photosynthesis at the expense of water loss via transpiration. Thus, quantification of the stomatal aperture has been instrumental to understanding plant environmental adaptation. However, quantifying the stomatal aperture is inherently time-consuming and cumbersome as it requires human labor to spot and measure stomatal pores in a leaf image captured by a microscope. To circumvent these limitations, various methods have been developed to facilitate the quantification of stomatal aperture in Arabidopsis thaliana, a model plant extensively used to study stomatal biology1,2,3,4,5,6. For instance, a porometer can be used to measure transpiration rate as a metric of stomatal conductance. However, this method does not provide direct information on the stomatal number and aperture that determine stomatal conductance. Some studies have used confocal microscopy techniques highlighting stomatal pores using a fluorescent actin marker, a fluorescent dye, or cell wall autofluorescence1,2,3,4,5. While these approaches facilitate the detection of stomata, the cost of both operating a confocal microscopy facility and preparing microscopy samples can be an obstacle to routine application. In a ground-breaking work by Sai et al., a deep neural network model was developed to automatically measure stomatal aperture from bright-field microscopic images of A. thaliana epidermal peels6. Yet, this innovation does not exempt researchers from the task of preparing an epidermal peel for microscopic observation. Recently, this obstacle was overcome by developing a portable imaging device that can observe stomata by pinching a leaf of A. thaliana, together with a deep learning-based image analysis pipeline that automatically measures stomatal aperture from leaf images captured by the device7.
Stomata contribute to plant innate immunity against bacterial pathogens. The key to this immune response is stomatal closure that restricts bacterial entry through the microscopic pore into the leaf interior, where bacterial pathogens proliferate and cause diseases8. Stomatal closure is induced upon recognition of microbe-associated molecular patterns (MAMPs), immunogenic molecules that are often common to a class of microbes, by plasma membrane-localized pattern recognition receptors (PRRs)9. A 22 amino acid epitope of bacterial flagellin known as flg22 is a typical MAMP that induces stomatal closure through its recognition by the PRR FLS210. As a countermeasure, bacterial pathogens such as Pseudomonas syringae pv. tomato DC3000 (Pto) and Xanthomonas campestris pv. vesicatoria have evolved virulence mechanisms to reopen stomata9,11,12. These stomatal responses to bacterial pathogens have been conventionally analyzed in assays in which either leaf epidermal peels, leaf discs, or detached leaves are floated on bacterial suspension, and then stomata are observed under a microscope followed by manual measurement of stomatal aperture. However, these assays are cumbersome and may not reflect stomatal responses to natural bacterial invasion that occur in a leaf attached to the plant.
Here, a simple method is presented to investigate stomatal closure and reopening during Pto invasion under the condition that closely mimics the natural plant-bacteria interaction. This method leverages the portable imaging device for direct observation of A. thaliana stomata on a leaf attached to the plant inoculated with Pto, together with the image analysis pipeline for automated measurement of stomatal aperture.
Protocol
1. Growing plants
- To break dormancy, resuspend A. thaliana (Col-0) seeds in deionized water and incubate them at 4 °C for 4 days in the dark.
- Sow the seeds on the soil and grow in a chamber equipped with white fluorescent light. Maintain the following growth conditions: temperature of 22 °C, light intensity of 6,000 lux (ca. 100 µmol/m2/s) for 10 h, and relative humidity of 60%.
- When needed, water the plants with a liquid fertilizer. Refrain from watering from 1 week to 2 days prior to inoculation and water well 1 day prior to inoculation.
2. Preparing bacterial inoculum
- Streak Pto from glycerol stock on solidified King's B (KB) medium (20 g of Tryptone, 1.5 g of K2HPO4, and 15 g of Glycerol for 1 L, 1.5% Agar) with 50 µg/mL rifampicin and incubate at 28 °C for 2 days.
- Inoculate a single colony to 5 mL of KB liquid medium with 50 µg/mL rifampicin and incubate at 28 °C with shaking at 200 rpm until late logarithmic growth phase.
- Centrifuge the culture at 6,000 x g for 2 min, discard the supernatant, and resuspend the pellet in 1 mL of sterile water. Repeat this step once again.
- Remove the supernatant, resuspend the pellet in 1 mL of stomata opening buffer (25 mM MES-KOH pH 6.15, 10 mM KCl), and measure the OD600.
- Dilute the suspension to OD600 0.2 with stomata opening buffer containing 0.04% silicone surfactant.
3. Spray inoculation of bacteria
- From 1 day before inoculation till the end of the experiment, expose plants to a light intensity of about 10,000 lux (ca. 170 µmol/m2/s).
- To ensure that most stomata are open, keep plants on a tray covered with a transparent lid under the light for at least 3 h before spray inoculation.
- Remove the lid and use an airbrush to spray the abaxial side of leaves with 2.5 mL of bacterial suspension per three plants on a single pot.
- Incubate the inoculated plants on a tray covered with a transparent lid to maintain a relative humidity of around 85%.
- Acquire images of stomata at 1 h and 3 h after spray inoculation using the method described in section 4.
4. Direct observation of stomata using the portable imaging device
NOTE: The portable stomatal imaging device is equipped with an LED light and a camera module and can acquire 2,592 × 1,944 (height × width; pixels) images with a resolution of about 0.5 µm/pixel.
- Connect the portable stomatal imaging device to a personal computer (PC) equipped with image acquisition software.
- Gently but completely remove water droplets from the inoculated leaves with a piece of paper.
- Open the top cover of the device, place the leaf on the stage, and close the top cover (Figure 1).
- Adjust the focus of the image by manipulating the adjuster screw, then click the Save Image button on the PC screen. The image will be acquired instantly. Typically, a focused image contains approximately 10 analyzable stomata. To obtain robust results, acquire stomatal images from six leaves of three different plants (two leaves per plant).
5. Manual measurement of stomatal aperture
NOTE: The ImageJ software can be downloaded at https://imagej.nih.gov/ij/download.html
- Open an image file in the ImageJ.
- Open the ROI manager by selecting Analyze > Tools > ROI Manager.
- Use the Straight-Line selection tool to draw a line corresponding to the width of a stoma (Figure 2) and register the ROI by clicking Add in the ROI Manager.
- Draw a line corresponding to the length of the same stoma (Figure 2A) and register the ROI as described in step 5.3.
- Click Measure in the ROI Manager to measure the width and length.
- Divide the width by the length to obtain the stomatal aperture (ratio). For robust quantification, use 60 or more stomata for each treatment and time point. Do not choose premature or obscure stomata for the analysis (Figure 2B, C).
6. Automated measurement of stomatal aperture
NOTE: The image analysis pipeline runs in Google Colaboratory, a cloud Python programming language executable environment. Users must have a valid Google account with a working Google Drive, Google Chrome browser, and a stable internet connection as a prerequisite.
- Download the Google Colaboratory notebook from Zenodo (https://doi.org/10.5281/zenodo.8062528), and open the notebook.
- Make a local copy of the notebook to Google Drive by selecting File > Save a copy in Drive. After a new tab appears, safely close the tab of the original notebook.
- Press the Execute button once below the Environmental Setup section in the notebook without unfolding the cell blocks to import the required libraries.
- Execute the Directory Settings section to create three folders used for analysis (e.g., example_result, inference_results, and model) in Google Drive.
NOTE: In this case, the folders named example_result, inference_results, and model are used as the parental directory, storing inference results and trained models, respectively. This notebook shows an example of directory construction as a representative procedure. To change the name, rewrite the pardir, infdir, or modeldir path. - According to the Preparation of the Images section, move the acquired images to example_result, grouped in image titles by treatment or sample (e.g., mock_1h_XXXXXX.jpg) for the final graph generation. Sample images are available from Zenodo (https://doi.org/10.5281/zenodo.8062528).
- Perform the Download Trained Models part to download the ONNX files of the trained models from Zenodo (https://doi.org/10.5281/zenodo.8062528) and place them under the model directory.
- Run the Inference and Measurement of Stomatal Aperture part to quantify the stomatal aperture from individual images. The result images with overlaid inference and the csv file named example_result.csv will be exported to the inference_results directory.
- Execute the Graph Generation section to create a graph about the stomatal aperture ratio, exported to the inference_results directory.
Representative Results
Following spray inoculation of Pto, stomata on leaves attached to the inoculated plants were directly observed by the portable stomatal imaging device. Using manual and automated measurements, the same leaf images were used to calculate stomatal aperture by taking ratios of width to length of approximately 60 stomata. Manual and automated measurements consistently indicated a decrease in the stomatal aperture in Pto-inoculated plants compared with mock-inoculated plants at 1 hour post inoculation (hpi) (Figure 3A,B), indicating that A thaliana plants close stomata in response to Pto invasion. At 3 hpi, the stomatal aperture in Pto-inoculated plants and mock-inoculated plants were virtually the same (Figure 3C,D), reminiscent of stomatal reopening by Pto. Remarkably, the automated measurement of stomatal aperture took only approximately 5 s to process one image (Table 1), reducing measurement time by more than 95% compared with the manual measurement. Thus, this protocol offers an operationally simple and labor-saving means to track the dynamic stomatal responses of A. thaliana to the bacterial pathogen.

Figure 1: Portable imaging device. Pictures depicting the portable imaging device with a leaf set on the stage (left) and with the top cover closed (right). Please click here to view a larger version of this figure.
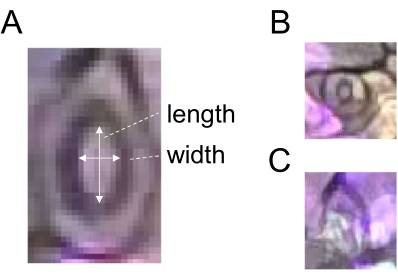
Figure 2: Schematic diagram of stomatal aperture measurement. (A) Stomatal aperture is determined by calculating the ratio of width to length of a stoma, as indicated by white arrows. (B) Premature and (C) obscure stomata should be excluded from the measurement. Please click here to view a larger version of this figure.
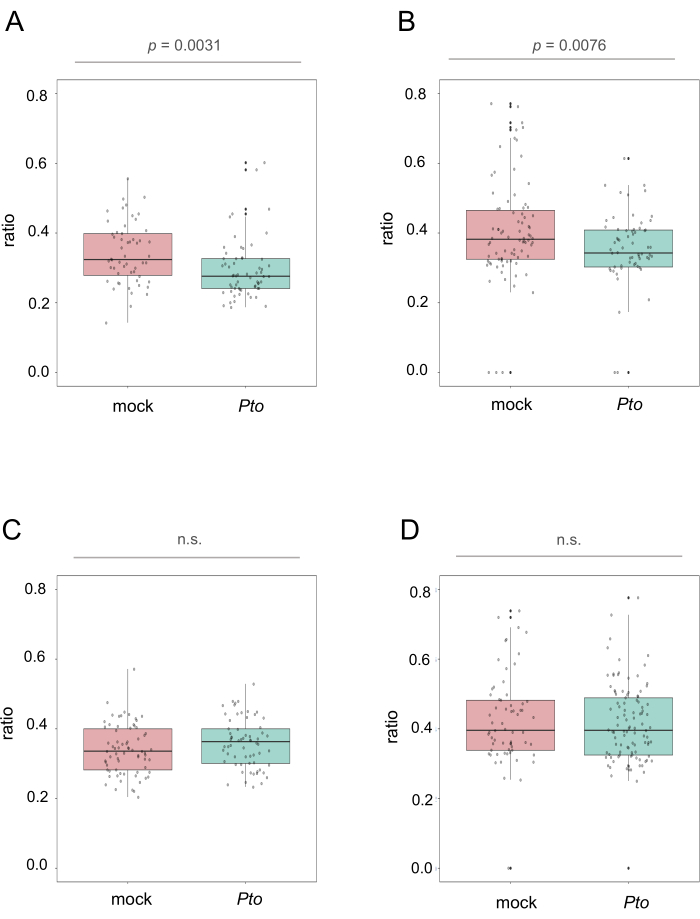
Figure 3: Stomatal responses to Pto in an intact whole plant. A. thaliana plants were spray-inoculated with mock or Pto, and stomata on leaves attached to the inoculated plants were directly observed at (A,B) 1 hpi and (C,D) 3 hpi by the portable stomatal imaging device. Stomatal aperture (ratio) was calculated by (A,C) manual and (B,D) automated measurements. P-values were calculated by a two-tailed t-test. Please click here to view a larger version of this figure.
| Processing time (s) | ||
| Method | Mean | SD |
| Manual | 130.1 | 48.8 |
| Automated | 4.7 | 0.8 |
Table 1: Processing time for manual and automated measurements of stomatal aperture per image. Means and standard deviations (SD) of processing time were calculated from the measurements of nine representative images.
Discussion
Previous studies used epidermal peels, leaf discs, or detached leaves to investigate stomatal responses to bacterial invasions9,11,12. In contrast, the method proposed in this study leverages the portable stomatal imaging device to directly observe stomata on a leaf attached to the plant after spray inoculation of Pto, mimicking natural conditions of bacterial invasion. In addition, because this method does not involve destructive sample preparation processes such as leaf detachment, leaf disc excision, and epidermal peeling, wounding and water loss associated with these sample preparation processes can be avoided. These effects should not be taken lightly, as wounding and water loss inevitably produce plant-derived signals such as the phytohormones jasmonate and abscisic acid that affect stomatal movements13,14.
There are several guidelines for the optimal use of the portable stomatal imaging device. Firstly, thoroughly removing water droplets from leaf surfaces is paramount to obtaining images of optimal clarity and focus. Secondly, it is recommended to take multiple images from identical leaf areas by manipulating the adjuster screw to fine-tune focus. This practice is expected to increase the number of analyzable stomata per leaf area, thereby mitigating potential sampling biases. Lastly, when pinching a leaf with the device, careful handling is required to avoid causing damage to the leaf. This is critical because wounding is one of the cues that elicit stomatal closure14.
Stomatal aperture tended to be more variable in the automated measurement than in the manual measurement (Figure 3). There are several possible reasons for this. It was previously reported that stomatal pores inferred by the image analysis pipeline often include cell walls and/or shadows of guard cells surrounding the stomatal pore7, which is not the case in manual measurement by human eyes. Stomata with unusual shapes may also affect the variation between manual and automated measurements, although the stomata detection model was trained to exclude such stomata from the analysis7. A few stomata were given zero values for stomatal aperture in the automated measurement but none in the manual measurement for unknown reasons. Future updates of the models may be necessary to address these issues. Nevertheless, as the automated stomatal aperture measurement essentially matched the manual measurement, the current version of the image analysis pipeline is of practical use.
The direct observation and automated measurement of stomatal aperture in A. thaliana described in this study hold promise for various applications toward elucidating the role of stomata in plant environmental adaptation. For instance, the presented method should be broadly applicable for rapidly quantifying stomatal aperture in an intact whole plant system after exposure to biotic stresses such as MAMPs and microbial pathogens as well as abiotic stresses such as drought. In support of this, a previous study successfully applied the image analysis pipeline to accurately quantify the stomatal aperture of "leaf discs" treated with the fungal toxin fusicoccin that induces stomatal opening or the stress hormone abscisic acid that induces stomatal closure7. Moreover, in principle, the portable imaging device allows for long-term time-course analysis of the stomatal aperture on a single identical leaf attached to the plant. This might shed light on new aspects of plant-microbe interactions because most studies have focused on stomatal responses to bacterial pathogens for the first several hours of the interaction9,10,11. It will also be interesting to employ and modify the presented method to explore stomatal responses to bacterial invasion under various environmental conditions. This is particularly relevant to understanding the impacts of environmental factors such as temperature, humidity, and soil water availability that affect stomatal movements and disease development by bacterial pathogens8,15. In conclusion, the presented method will be envisioned to accelerate research on stomatal functions in and beyond plant-microbe interactions under hitherto unattainable experimental settings.
Disclosures
The authors have nothing to disclose.
Acknowledgements
We thank all the members of the research project, 'Co-creation of plant adaptive traits via assembly of plant-microbe holobiont', for fruitful discussions. This work was supported by Grant-in-Aid for Transformative Research Areas (21H05151 and 21H05149 to A.M. and 21H05152 to Y.T.) and Grant-in-Aid for Challenging Exploratory Research (22K19178 to A. M.).
Materials
| Agar | Nakarai tesque | 01028-85 | |
| Airbrush kits | ANEST IWATA | MX2900 | Accessory kits for SPRINT JET |
| Biotron | Nippon Medical & Chemical Instruments | LPH-411S | Plant Growth Chamber with white fluorescent light |
| Glycerol | Wako | 072-00626 | |
| Half tray | Sakata | 72000113 | A set of tray and lid |
| Hyponex | Hyponex | No catalogue number available | Dilute the solution of Hyponex at a ratio of 1:2000 in deionized water for watering plants |
| Image J | Natinal Institute of Health | Download at https://imagej.nih.gov/ij/download.html | Used for manual measurement of stomatal aperture |
| K2HPO4 | Wako | 164-04295 | |
| KCl | Wako | 163-03545 | |
| KOH | Wako | 168-21815 | For MES-KOH |
| MES | Wako | 343-01621 | For MES-KOH |
| Portable stomatal imaging device | Phytometrics | Order at https://www.phytometrics.jp/ | Takagi et al.(2023) doi: 10.1093/pcp/pcad018. |
| Rifampicin | Wako | 185-01003 | Dissolve in DMSO |
| Silwet-L77 | Bio medical science | BMS-SL7755 | silicone surfactant used in spray inoculation |
| SPRINT JET | ANEST IWATA | IS-800 | Airbrush used for spray inoculation |
| SuperMix A | Sakata seed | 72000083 | Mix with Vermiculite G20 in equal proportions for preparing soil |
| Tryptone | Nakarai tesque | 35640-95 | |
| Vermiculite G20 | Nittai | No catalogue number available | Mix with Super Mix A in equal proportions for preparing soil |
| White fluorescent light | NEC | FHF32EX-N-HX-S | Used for Biotron |
References
- Shimono, M., Higaki, T., Kaku, H., Shibuya, N., Hasezawa, S., Day, B. Quantitative evaluation of stomatal cytoskeletal patterns during the activation of immune signaling in Arabidopsis thaliana. PLoS One. 11, e0159291 (2016).
- Bourdais, G., et al. The use of quantitative imaging to investigate regulators of membrane trafficking in Arabidopsis stomatal closure. Traffic. 20 (2), 168-180 (2019).
- Higaki, T., Kutsuna, N., Hasezawa, S. CARTA-based semi-automatic detection of stomatal regions on an Arabidopsis cotyledon surface. Plant Morphology. 26 (1), 9-12 (2014).
- Eisele, J. F., Fäßler, F., Bürgel, F., Chaban, C. A. A rapid and simple method for microscopy-based stomata analyses. PLoS One. 11, e0164576 (2016).
- Chitraker, R., Melotto, M. Assessing stomatal response to live bacterial cells using whole leaf imaging. Journal of Visualized Experiments. 44, 2185 (2010).
- Sai, N., et al. StomaAI: an efficient and user-friendly tool for measurement of stomatal pores and density using deep computer vision. New Phytologist. 238 (2), 904-915 (2023).
- Takagi, M., et al. Image-based quantification of Arabidopsis thaliana stomatal aperture from leaf images. Plant and Cell Physiology. pcad018, (2023).
- Melotto, M., Zhang, L., Oblessuc, P. R., He, S. Y. Stomatal defense a decade later. Plant Physiology. 174 (2), 561-571 (2017).
- Melotto, M., Underwood, W., Koczan, J., Nomura, K., He, S. Y. Plant stomata function in innate immunity against bacterial invasion. Cell. 126 (5), 969-980 (2006).
- Zeng, W., He, S. A prominent role of the flagellin receptor FLAGELLIN-SENSING2 in mediating stomatal response to Pseudomonas syringae pv tomato DC3000 in Arabidopsis. Plant Physiology. 153 (3), 1188-1198 (2010).
- Zheng, X. Y., et al. Coronatine promotes Pseudomonas syringae virulence in plants by activating a signaling cascade that inhibits salicylic acid accumulation. Cell Host and Microbe. 11 (6), 587-596 (2012).
- Raffeiner, M., et al. The Xanthomonas type-III effector XopS stabilizes CaWRKY40a to regulate defense responses and stomatal immunity in pepper (Capsicum annuum). The Plant Cell. 34 (5), 1684-1708 (2022).
- Munemasa, S., Hauser, F., Park, J., Waadt, R., Brandt, B., Schroeder, J. I. Mechanisms of abscisic acid-mediated control of stomatal aperture. Current Opinion in Plant Biology. 28, 154-162 (2015).
- Förster, S., et al. Wounding-induced stomatal closure requires jasmonate-mediated activation of GORK K+ channels by a Ca2+ sensor-kinase CBL1-CIPK5 complex. Developmental Cell. 48 (1), 87-99 (2018).
- Cheng, Y. T., Zhang, L., He, S. Y. Plant-microbe interactions facing environmental challenge. Cell Host and Microbe. 26 (2), 183-192 (2019).
