Bioluminescence recording was conducted to assess the circadian rhythmicity of human intestinal enteroids (HIEs) under two distinct conditions: stem cell-enriched conditions using intestinal organoid growth medium (Figure 3) versus differentiation-inducing conditions, which was achieved by replacing the intestinal organoid growth medium with a differentiation medium. On the day of the experiment, we synchronized the circadian clocks by performing a 1 h treatment with 100 nM Dexamethasone. Subsequently, we replaced the medium with either 3 mL of intestinal organoid growth medium or differentiation medium supplemented with 200 µM D-Luciferin and initiated the monitoring of bioluminescence activity. Following a recording period of 4-5 days, we conducted a Fast Fourier Transform (FFT) analysis to determine the period of the observed circadian rhythms. Figure 3 shows the temporal dynamics of Bmal1 promoter activity in the aforementioned conditions. Under differentiating conditions, Bmal1-luc bioluminescence exhibited robust circadian oscillations with a mean periodicity of 22.92 h. In contrast, HIEs demonstrated disrupted circadian rhythms in stem cell-enriched conditions (Figure 3). These data show that HIEs in stem cell-enriched conditions do not show robust circadian oscillations. In other words, one would need to assess the impact of growth conditions with respect to circadian rhythms before investigating circadian clock-dependent functions in organoids.
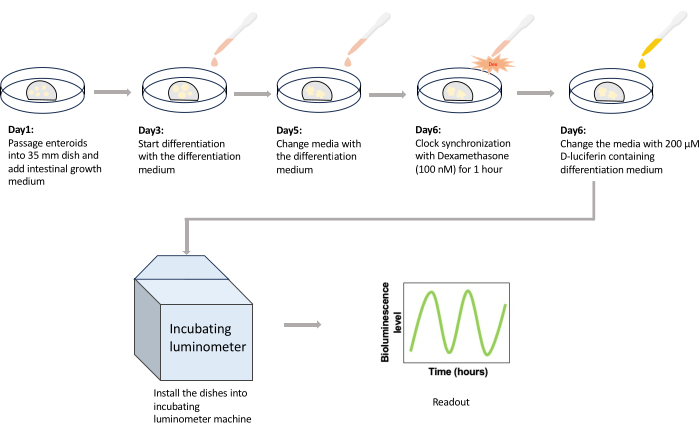
Figure 1: Schematic drawing of the protocol. HIEs are passaged into 35 mm dishes on day 1 and cultured with 2 mL of growth medium until day 3. On the third day after passage, the medium is replaced with 2 mL of prewarmed differentiation medium to initiate the differentiation of enteroids. Medium change is repeated with 2 mL of prewarmed medium on day 5 to prevent nutrition depletion. On day 6, enteroids are treated with 100 nM Dexamethasone for 1 h for circadian clock synchronization. After an hour, the media is replaced with 3 mL of 200 µM D-luciferin-containing medium, and the dishes are installed in the incubating luminometer. After recording the bioluminescence expression for 4-5 days, the experiment is terminated, and generated data are exported as spreadsheets for further analysis. Abbreviation: HIE = human intestinal enteroid. Please click here to view a larger version of this figure.
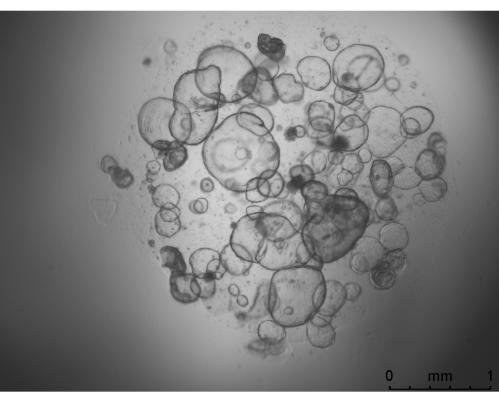
Figure 2: A representative image for HIEs under growth medium. The image was taken by a widefield microscope with a 25x objective. Scale bar = 1 mm. Abbreviation: HIE = human intestinal enteroid. Please click here to view a larger version of this figure.
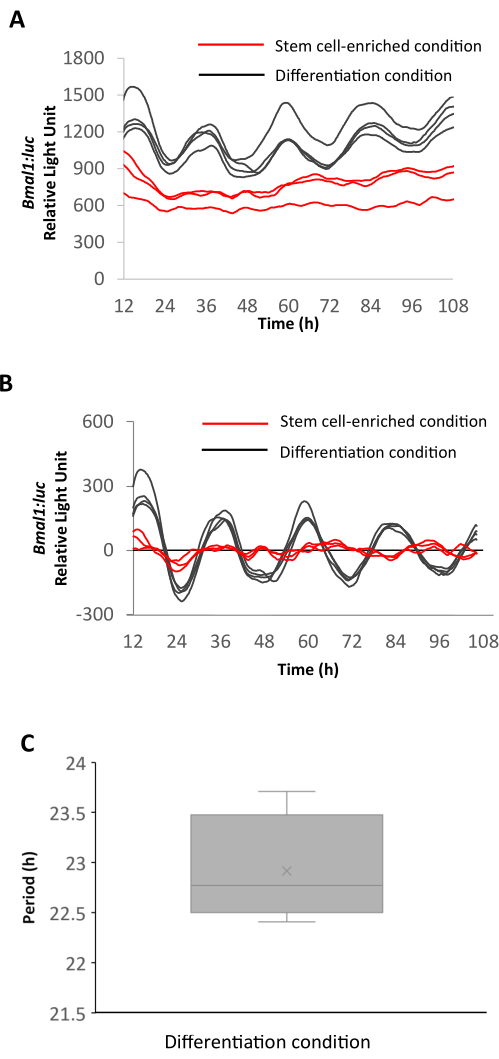
Figure 3: Bioluminescence recordings from HIEs from stem cell-enriched and differentiation conditions. (A) Raw and (B) detrended bioluminescence recording data of Bmal1-luc HIEs under stem cell-enriched (red) and differentiation (black) conditions. (C) Period of Bmal1-luc oscillations in HIEs under differentiation conditions. The period is calculated with FFT analysis; n = 3 for Bmal1-luc HIEs under stem cell-enriched condition and n = 4 for Bmal1-luc HIEs under differentiation condition. Error bars indicate SD. Abbreviations: HIE = human intestinal enteroid; FFT = Fast Fourier Transform. Please click here to view a larger version of this figure.
| Differentiation Media Recipes | ||||
| Name of Reagent | Company | Catalog Number | Final Concentration | Comments |
| Advanced DMEM/F12 | Life Technologies | 12634-028 | Add as the last component. If you prepare 50 mL media, after adding all other components, fill the rest up to 50 mL with Advanced DMEM/F12 at the end. | |
| A 83-01 | Sigma Aldrich | SML0788 | 500 nM | |
| B-27 Supplement (50x) | Gibco | 17504-044 | 1x | |
| Gastrin I Human | Sigma Aldrich | G9020 | 10 nM | |
| GlutaMAX | Gibco | 35050061 | 1x | |
| HEPES | Gibco | 15630080 | 10 mM | |
| N-2 Supplement (100x) | Gibco | 17502-048 | 1x | |
| N-Acetyl-L-cysteine | Sigma Aldrich | A9165 | 1 mM | |
| NOGGIN Conditioned Medium | 5% | Made in the lab | ||
| Penicillin-Streptomycin | Gibco | 15140122 | 100 U/mL | |
| Recombinant murine EGF | PeproTech | 315-09 | 50 ng/mL | |
| R-SPONDIN Conditioned Medium | 10% | Made in the lab |
Table 1: Composition of HIE differentiation media23.
