The design of the enzymatic microelectrode biosensor allows the real-time detection of analytes in whole kidneys. The general experiment design for either ex vivo or in vivo studies is illustrated in Figure 1.The sensors used and the surgical procedures differ depending on whether the study is ex vivo or in vivo.
To obtain reproducible results, accurate pre- and post- calibrations are critical. Figure 6A shows a representative trace of the signal seen from the ex vivo ATP sensor during calibrations. Note that apyrase quickly eliminated the ATP signal. Catalase had no effect on the ATP signal and demonstrates its specificity to the H2O2 it creates (Figure 6B). The calibration procedure produces a linear fit that is used to calculate the dynamic changes of ATP (Figure 6A, right panel). In vivo sensor calibration produces a similar trace to that of the ex vivo sensor. However, this sensor detects reductive instead of oxidative currents and as such the current produced is negative. Calibration of these sensors also produces a linear fit in a 0.3 to 80 µM range (Figure 7A right panel). Specificity of in vivo sensor to ATP over other purine products is shown in Figure 7B.
The approach described here allows us to measure both basal levels of endogenous substances and acute changes in response to drug infusion12. Shown in Figure 11 are Ang II-induced changes of interstitial endogenous concentration of H2O2 in salt-resistant and salt-sensitive rats. For these experiments, freshly isolated kidneys from Sprague Dawley (SD) or Dahl salt-sensitive (SS) rats were perfused with 1 µM Ang II under constant laminar flow (650 µl/min). As shown in Figure 11, Ang II induces the acute release of H2O2 in kidneys from both SD and SS rats. However, the maximum amplitude of each response was significantly elevated in SS rats, especially when the animals were fed with a high salt diet12. Shown in Figure 12 is a representative application of the biosensors in vivo. The infusion of catalase (5 µg/ml) completely blocks the H2O2 signal detected by the biosensor. These experiments illustrate the use of specific enzymatic biosensors combined with the amperometric technique for the detection of basal levels of endogenous substances and real-time measurements of their release in response to pharmacological intervention. Further applications of this approach to study the role of purinergic signaling and hydrogen peroxide will improve our knowledge and understanding of renal pathologies like salt-sensitive hypertension, renal oxidative stress and chronic kidney disease.
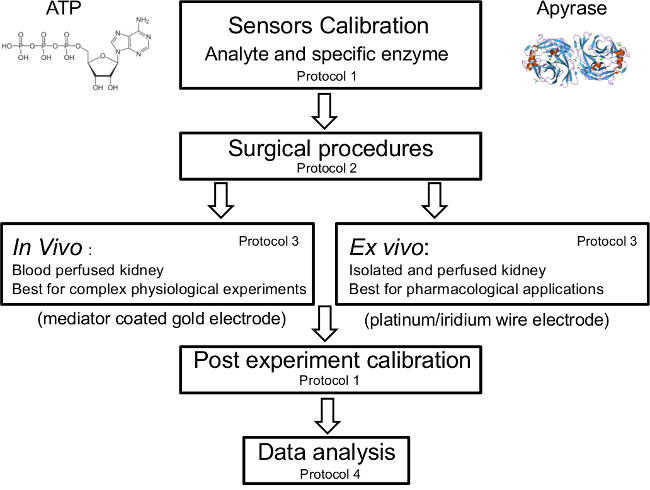
Figure 1. Schematic of the Protocols. Calibration with analyte of interest and testing its specificity is done immediately prior to the start of the experiments. The in vivo protocol should be followed for complex physiological experiments and ex vivo for pharmacological applications. Post experiment calibration should be done and taken into account during the data analysis. Please click here to view a larger version of this figure.

Figure 2. Enzymatic Microelectrode Sensor. The two sensor sizes used for ex vivo applications are shown, 50 µm and 125 µm diameters. Each sensor wire extends into a capillary tube to connect to a gold end connector (not shown). Inset shows a sensor tip coated with enzymes for ATP detection. Please click here to view a larger version of this figure.

Figure 3. Dual Channel Potentiostat. The potentiostat channels labeled CH1 and CH2. 1) CH1 the ATP sensor connection and 2) CH2 the Null sensor connection 3) connections that are electrically coupled to the reference electrode. Please click here to view a larger version of this figure.

Figure 4. Sensor Setup. The data acquisition is performed in a Faraday cage to reduce electrical noise and on a high-performance lab table for a vibration-free working surface. 1) A temperature-controlled surgical table is used to maintain animal body temperature during physiological experiments 2) the micromanipulators are attached to magnetic mounting adaptors for flexible positioning of the sensors during experiment 3) A light source is needed for surgical procedures and sensor insertion 4) dual channel potentiostat 5) holder for the sensors. Please click here to view a larger version of this figure.
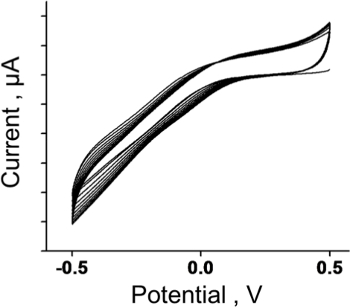
Figure 5. Cyclic Voltammetry. For ex vivo studies, sensor cyclic voltammetry is conducted for 10 cycles between -0.5 and 0.5 V prior to the calibration protocol. Please click here to view a larger version of this figure.
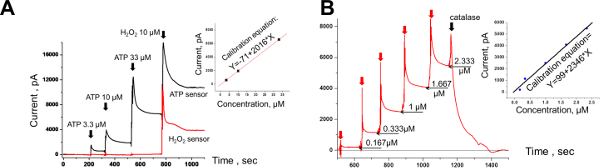
Figure 6. Amperometry Calibration for Ex Vivo. (A) Calibration uses the addition of known ATP concentrations to the bath solution. Corresponding amperometric values recorded at the asymptote level (black trace). The currents obtained create a calibration equation. An example is shown on the right panel. The red trace is the current of the Null sensor which responds to only the addition of H2O2. (B) The Null sensor is calibrated with the addition of known H2O2 concentrations (red arrows) to the bath solution and the asymptotes amperometric values are recorded (thin black arrows). Addition of catalase to the bath solution (thick black arrow) results in rapid current decay. The right panel shows the Null sensor calibration equation. Please click here to view a larger version of this figure. Please click here to view a larger version of this figure.
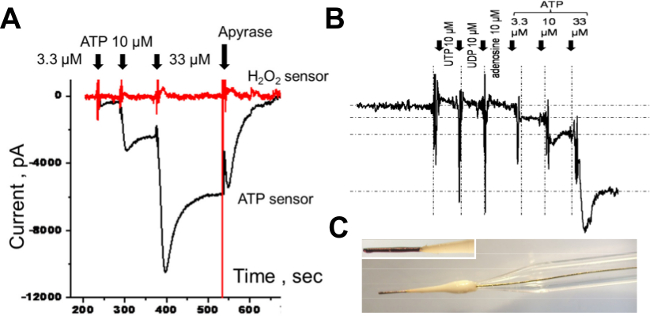
Figure 7. Amperometry Calibration for In Vivo. Calibration of the in vivo electrodes is performed in a similar fashion to that detailed in the ex vivo studies except reduction reactions cause a reverse current (polarity). (A) The addition of known ATP concentrations produces an amperometric current on the ATP sensor (black trace) but has no effect on the Null sensor (red trace). Addition of apyrase extinguishes the current of the ATP sensor. (B) The specificity of the ATP sensor is confirmed by the addition of 10 µM of different purinergic agents (UTP, UDP and adenosine). Further applications of ATP provide a stable detectable amperometric current. (C) Microphotograph of the in vivo sensors based on a mediator coated gold electrode. Please click here to view a larger version of this figure.
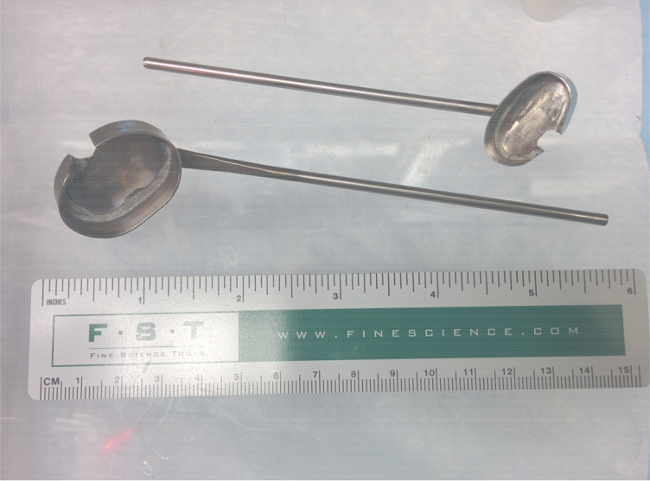
Figure 8. Kidney Cups. In the in vivo studies, kidneys are held still using the stainless steel kidney cups shown. Two sizes of cups are used to accommodate kidney size variation. These cups reduce the movement artifacts which are generated by the animal’s respiration. Please click here to view a larger version of this figure.

Figure 9. Ex Vivo Isolated and Perfused Kidney. The isolated kidney is placed into a 1) Petri dish coated with a thick silicone bottom for pin insertion 2) the renal artery is cannulated and attached to a syringe pump for constant perfusion during experiment 3) the ATP sensor, 4) and Null sensor are inserted into the kidney 5) the reference electrode is placed near the kidney submerged into the bath solution. Please click here to view a larger version of this figure.
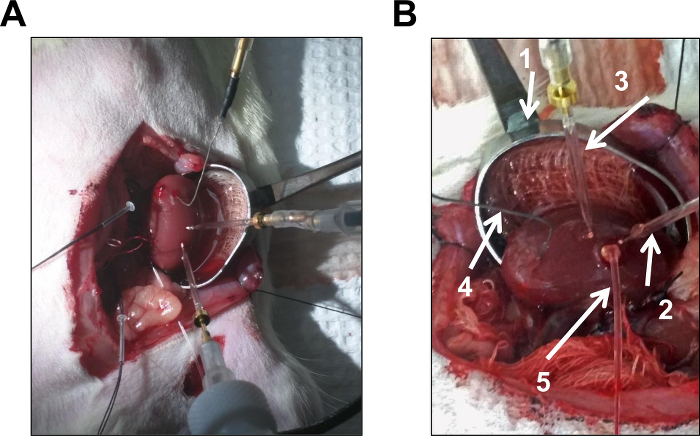
Figure 10. In Vivo Blood Perfused Kidney. (A) The left kidney is exposed and placed in a kidney cup, the right kidney remains intact inside the animal. Both sensors are inserted into the kidney. BSA:NaCl is infused via the catheterized jugular vein to counteract the fluid loss caused by the large mid-line incision (B) Example of the in vivo experiment with an implanted interstitial catheter for direct pharmacological applications 1) the kidney is held by a kidney cup 2) ATP sensor 3) Null sensor and 4) reference electrode are inserted into the ventral surface of the kidney 5) catheter implanted into the kidney and attached to a peristaltic pump for laminar pharmacological infusions. Please click here to view a larger version of this figure. Please click here to view a larger version of this figure.
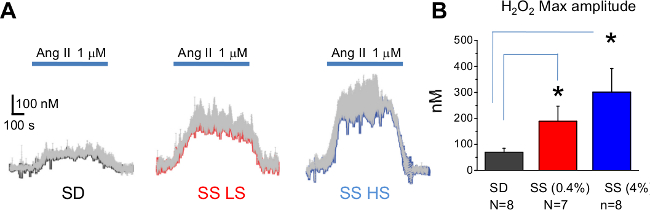
Figure 11. Ex Vivo Analysis of H2O2 in the kidney. Ang II perfusion causes H2O2 release in the rat kidney cortex. (A) Real time changes of the mean H2O2 concentration (gray bars show standard error) from a total of N = 8 applications (4 kidneys from 4 different rats). Bars on the top represent Ang II application. (B) The maximal H2O2 concentration amplitude values during Ang II perfusion for Sprague Dawley (SD) and Dahl salt-sensitive (SS) rats fed a low and high salt diets, respectively. * – P < 0.05 versus SD rats. The figure is adapted from reference12 with permission. Please click here to view a larger version of this figure.
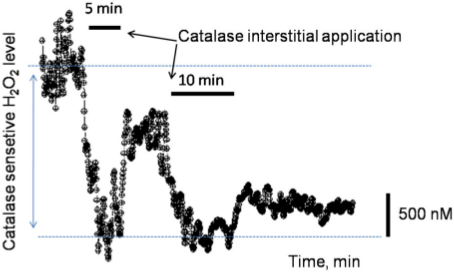
Figure 12. In Vivo Analysis of H2O2. Example of the in vivo assessment of interstitial H2O2 concentration in the medulla of a SS rat fed a low salt diet (as shown in Figure 10B). The interstitial application of catalase via an implanted catheter for the 5 min interval produced a complete blockade of the H2O2 signal in the renal medulla. Reduction of catalase, from washout by renal blood flow, resulted in a partial recovery of the signal, which was blocked again by an additional catalase application (10 min). Please click here to view a larger version of this figure.
