In humans, responses to infection, tissue inflammation, cellular recruitment, tissue remodeling and the regulation of tissue homeostasis are complex events involving different cell types. Hence, these processes are best studied in the local tissue environment. Previously, this has mainly been possible using experimental animal models. However, the widely used experimental animals hold many limits as they often respond to pathogens in a different way than humans and also display a different course of disease 1. A human in vitro lung tissue model holds the possibilities to study specific immune responses in the human lung.
Human tuberculosis infection (TB) is mainly a disease affecting the lungs. Mycobacterium tuberculosis (M. tuberculosis), the causative agent of TB, reaches the lung via aerosol droplets that are transported to the alveolar space, where the bacteria are engulfed by pulmonary dendritic cells and alveolar macrophages as part of the innate immune response to the infection 2,3. Phagocytosis of the pathogen leads to the compartmentalization of the bug within a phagosome and ideally results in the neutralization and killing of the pathogen by the phagocyte. Up to 50% of individuals exposed to M. tuberculosis are believed to be able to clear the infection through the innate immune response 4. Other outcomes of infection are clearance by the adaptive immune system at a later stage, latent infection or in worst cases chronic active disease 5.
Previously there have been no in vitro tissue models for studies of human TB. Single cell cultures of human macrophages or other peripheral blood cells have often been used 6,7. The disadvantage of this approach is that they cannot reflect the dynamics of different cell types operating together in a lung tissue exposed to M. tuberculosis. Thus, there is a need for an in vitro model to be able to perform functional and mechanistic studies on TB. The cell-based in vitro human lung tissue model described here was originally established by our group for studies on dendritic cell functions 8. We have adapted this method for the study of TB.
The human lung tissue model presented here consists of tissue-specific epithelial cells and fibroblasts 8. These cells are cultured in a matrix of collagen on top of a porous membrane in a transwell insert and form structures resembling normal human lung tissue (Figure 1). When exposed to air the cells start to secrete mucus at the apical side 8. By implanting human primary macrophages infected with M. tuberculosis to the model, we have observed how the immune cells migrate in the tissue and form early stages of TB granulomas 9. This is the first human tissue model described for TB and it poses a promising tool for studying innate immune responses to TB and other diseases of the lung. To date, we have used only monocytes and macrophages as immune cells in the model but the level of complexity can be increased by inclusion of additional relevant cell types.

Figure 1. Schematic outline of the lung tissue model. (A) The model is composed of human lung-specific epithelial cells, M. tuberculosis-infected primary macrophages and red dye labeled monocytes seeded onto collagen embedded fibroblasts prepared on a transwell filter. Exposure of the tissue model to air initiates production of extra-cellular matrix proteins, mucus secretion and stratification by the epithelium. The 3D tissue model thus developed is a useful tool to study M. tuberculosis infection in an environment that closely resembles a human lung. (B) Representative microscopic images of the different steps in the preparation of the tissue model. (C) Complete structure of the lung model tissue section. Scale – 100 µm. Please click here to view a larger version of this figure.
A 3D lung tissue model for human TB can be effectively used to study the host-pathogen interactions in M. tuberculosis infection. The basic steps of this method, representative microscopic images of different steps and an overall microscopic structure of a tissue section are outlined in Figure 1. The model has several components of human lung tissue, including lung fibroblasts, bronchial epithelial cells and primary monocytes/macrophages embedded in the 3D tissue environment. Besides incorporating components of human lung tissue, the model resembles physiological conditions namely stratification of epithelia and mucus secretion.
An example for the use of the lung tissue model in monitoring a TB infection is presented in Figure 2. For visualizing the M. tuberculosis-immune cell migration and interaction, we introduced macrophages infected with M. tuberculosis that express GFP (green) together with the freshly isolated PKH26-labelled monocytes (red) into the tissue model (blue, DAPI stained for nuclei). On day 7 post addition of M. tuberculosis-infected cells to the tissue model, confocal microscopy reveals clustering of red monocytes at the site of infection (green) (Figure 2), which mimics the hallmark lesions of human TB 9.
A series of representative images for 3D visualization of M. tuberculosis-infected tissue model and quantification of cell clusters is shown in Figure 3. The 3D visualization gives the user flexibility to interact, examine and quantify several features in a 3D image. The spatial arrangement of green bacteria and red monocyte clusters can be seen from the apical, rotational and lateral view as illustrated in Figure 3B, which reveals clustering of monocytes at the site of M. tuberculosis. The clusters were not observed in uninfected tissues (Figure 3A). We quantified the size and number of monocyte cell clusters and found that the size (volume) of cell clusters is enhanced (p<0.001), while the number of individual monocytes decreased (p<0.01) in M. tuberculosis infected tissues as compared to uninfected tissue models (Figure 3C and 3D). This data validates our previous finding of early granuloma formation in M. tuberculosis infection observed in lung tissue models analyzed in 2D tissue sections 9.
Our data suggests that the tissue model provides a natural 3D habitat to investigate the complex host cell-M. tuberculosis communication network. We also found that 3D visualization and quantitative analysis are better tools for studying the features in the tissue model (Figure 3). Quantification of a cell cluster (granuloma for instance) often stretches to several cell layers and can be completely captured by a 3D quantitative analysis. Moreover, visualization of exact spatial and temporal features of individual cells or bacteria in the model allow live-imaging, migration and tracking studies in a designated laboratory.
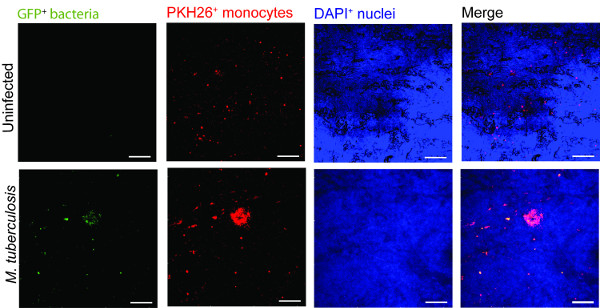
Figure 2. Monocyte in the tissue model cluster around virulent M. tuberculosis. Representative confocal images of uninfected and M. tuberculosis infected tissue model is presented. Panels from green (M. tuberculosis-GFP), red (PKH26-labelled monocytes), blue (DAPI-stained nuclei) and merged channels show the recruitment of monocytes in the infected tissue as compared to uninfected tissues. Scale – 100 µm. Please click here to view a larger version of this figure.
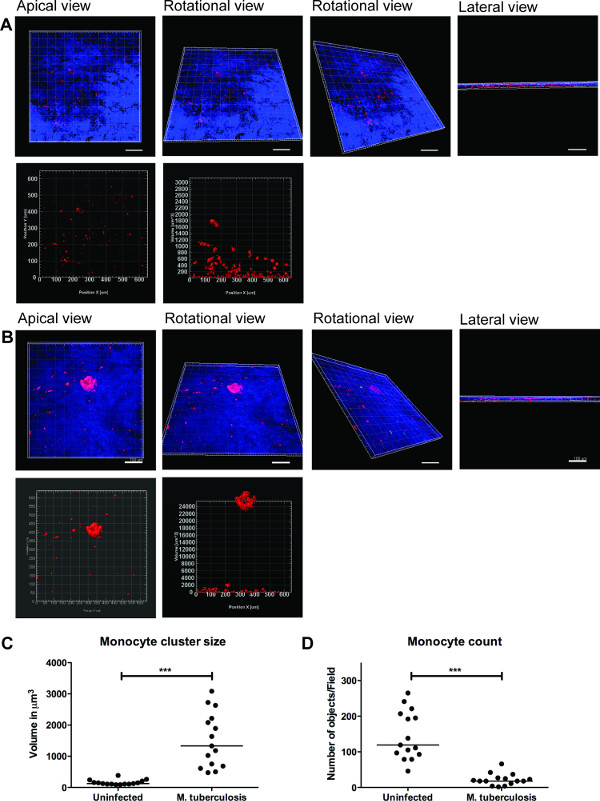
Figure 3. 3D visualization and quantitative analysis of tissue model provide useful information. Representative images of 3D visualization of the entire tissue model (A) uninfected tissue, (B) infected with M. tuberculosis, through optical sectioning using Zeiss LSM700 confocal microscope and quantitative analysis by Imaris image processing software (version 7.6.8). These images were acquired at 20X magnification, 14 z-stacks covering a tissue thickness of 19.5 µm with 1.5 µm interval, allowing the visualization from apical, rotational horizontal, rotational vertical and lateral view (A and B). (C) Quantitative analysis of monocyte cell clusters reveal enhanced (p<0.0001) size of early granuloma clusters after M. tuberculosis infection when compared to absence of infection. (D) Quantification of number of monocytes showed a decline (p<0.01) in infected tissue as compared to uninfected tissue, reiterating more clusters in the infected tissue. Green –M. tuberculosis-GFP, Red – PKH26-labelled monocytes, Blue – cell nuclei, Scale – 100 µm. Please click here to view a larger version of this figure.
