While 3D assays can be done within the same stiffness of collagen gel, varying the gel stiffness can be used to determine how the cells will respond to mechanical changes in their cellular microenvironment. A stiff collagen hydrogel is defined as a gel where the embedded cells are unable to locally contract the surrounding collagen. The intrinsic contractility of different cell types is unique, and thus it is best to begin with a simple contractility curve to establish the collagen concentration that will be sensed as compliant and stiff.
Begin by holding the cell numbers constant and increasing the concentration of the collagen gel. The stiffness of the gel will scale exponentially with concentration, as shown previously10,20. A representative example of a contractility assay is shown in Figure 1. Here, 50,000 4T1 cells are embedded in either a 2 mg/ml or 3.5 mg/ml collagen gel for 5 days. The 4T1 cell line is a highly contractile, metastatic cancer line, but is only able to contract the high density gel by approximately 10% (Figure 1a, c). This low level of contraction is typical for a stiff gel classification. Comparable low density gels (2 mg/ml, Figure 1b, c) are contracted 57% (n = 4) during the same time frame. It is important to note that increasing the amount of collagen ligand can impact cell signaling pathways, therefore a second means to vary the stiffness is to hold the collagen concentration constant and compare gels that are restrained (stiff) by being left attached to the bottom of the dish or contractile (compliant) because they have been released to float freely in the cell medium. This can serve as a good control to determine whether changes in cellular signaling result from increased collagen ligand or the stiffness of the matrix.
While collagen gel contraction assays are straightforward, there are a few items to be aware of when using these types of assays. For one, under high density conditions it is important to verify that there is not a loss of cell viability. We have observed that there is a ceiling for the stiffness of the microenvironment that a cell will tolerate, and have observed increased apoptosis or cell death in some cell lines, which can cause subsequent results to be misinterpreted. There should be no appreciable loss of cell viability between high and low stiffness conditions. This can be determined by Western blotting for caspase activity or some other assay for measuring cell viability. Secondly, collagen gels can also be contracted too much thereby creating a stiffer environment, and diminishing the differences between high and low stiffness conditions. We recommend not exceeding contractions of 60 – 70%. To maximize reproducibility, it is important to optimize the timeframe of the experiment, the initial cell seeding density, and the stiffness of the matrix to the contractility of the cell type in use.
Once mastery of embedding cells within 3D matrices is achieved, assays can be modified to alter the organization and orientation of the collagen fibers. This is achieved by the use of PDMS microfluidic channels (Figure 2). For the purpose of this manuscript, two similar conformations were used, a narrow and a wide channel, with 3 openings or ports (Figure 2b). In a typical experiment to determine the cellular response to random compared to aligned matrices, unpolymerized collagen is drawn through the microfluidic channel from port A to port C with cells being added to port B. The degree of collagen alignment is modulated by the rate of collagen flow such that higher collagen flow rates yield better alignment. The use of narrower channels coupled with higher vacuum pressures enhances the alignment of the collagen network, while a wider channel used in conjunction with low vacuum pressures generates random matrices. The organization of the fibrillar network can be visualized by second harmonic generation, as shown in representative images (Figure 2c, d). In these images, the collagen fibers (indicated by the arrowheads and pseudocolored in purple) align more consistently with the axis of collagen flow in the narrow channels (Figure 2d). The collagen fibers in wide channels have a variety of orientations, and have a more random distribution (Figure 2c).
The difference between wide and narrow channels is discernable to the human eye, but can also be analyzed via software developed specifically for fiber analysis. CT-FIRE, an open source and freely available software, utilizes a curvelet-based algorithm for pre-processing followed by a fiber extraction algorithm to quantify fiber angle, thickness, length, and straightness21. For each condition, more than 500 images were randomly sampled from different locations within the channels, and more than 125,000 fibers were extracted and analyzed by CT-FIRE. The subsequent fiber angle histogram generated for the narrow channels (Figure 2f) shows a clear enrichment of fibers oriented parallel to the direction of flow. In contrast, wide channels have a more equivalent fiber angle distribution (Figure 2e) which is consistent with a more random matrix.
Importantly, the use of microfluidic devices, as a technique to control the organization and orientation of collagen fibers, allows for assessment of various cellular responses to altered matrix conditions. For example, Sung et al. determined that viability of human mammary fibrobasts is dependent on collagen fiber.15 Additionally, recent work from our laboratory demonstrates that breast carcinoma cells migrate more persistently on aligned collagen fibers compared to random collagen matrices.3 These results help to better define the role of the microenvironment within the context of mammary biology and breast cancer. Moving forward, the ability to precisely control the 3D cellular microenvironment will allow for continued investigation of important cellular responses within the context of many different disease states.
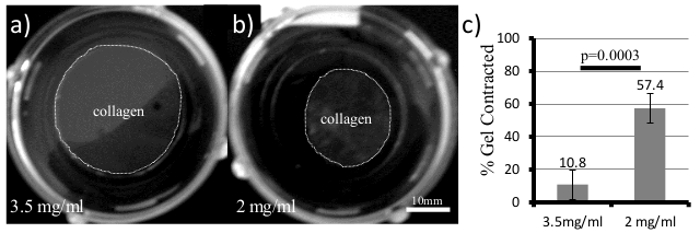
Figure 1. 3D Collagen Gels for Measuring Cell Contractility and Cellular Responses to ECM Stiffness. 4T1 cells were plated in both (a) 3.5 mg/ml and (b) 2 mg/ml collagen gels and allowed to grow for 5 days. Over time, the collagen gel (Gray area within dotted line) will contract and the diameter is measured and quantified in (c, Student's t-test, error bars +/- SD). Please click here to view a larger version of this figure.
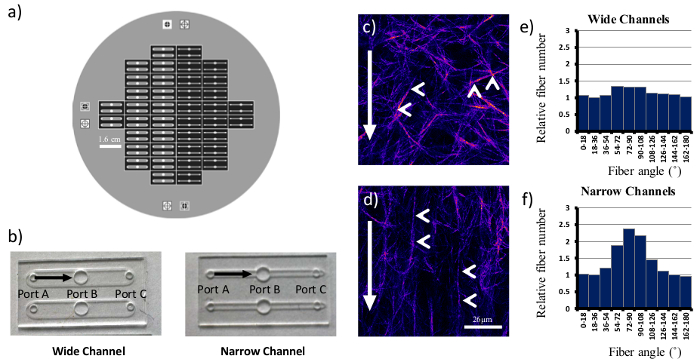
Figure 2. Generating Random and Aligned Collagen Matrices Using PDMS Microchannels. A six inch silicon wafer (a) was used as a template for both wide and narrow PDMS microchannels (b). Identical collagen concentrations were drawn through both types of channels, and individual fibers (arrowheads, c, d) align with the direction of flow (arrows) in narrow microchannels (d). Wide microchannels yielded more random matrices. Fiber angles with respect to the bottom of image were counted and shown in (e) and (f). Scale bar = 26 µm. Please click here to view a larger version of this figure.
