To investigate whether an Aβ1-42 monomer can form an Aβ1-42 oligomer after preparation, TEM analysis was used. No visible aggregates were observed in the HFIP-dissolved Aβ1-42 monomer sample (Figure 1A). Moreover, mainly globular aggregates with a diameter of around 10 – 80 nm were observed in the Aβ1-42 sample after 48 h of shaking, suggesting that Aβ1-42 forms oligomers after preparation (Figure 1B).

Figure 1. TEM analysis of Aβ1-42 monomer and Aβ1-42 oligomer-rich samples. The HFIP-dissolved Aβ1-42 monomer sample (10 µM) and the Aβ1-42 oligomer-rich sample (10 µM) prepared according to this protocol were examined by TEM. The scale bar = 200 nm. Please click here to view a larger version of this figure.
Additionally, dot blotting analysis was used to evaluate the relative amounts of Aβ1-42 oligomer in the samples. An A11 antibody could react with a subset of toxic Aβ oligomer with anti-parallel β-sheet structures. An OC antibody reacts with fibrillar aggregates with parallel in-register β-sheet structures. By using these antibodies, we demonstrated that A11-positive Aβ1-42 oligomers, but not OC-positive Aβ1-42 fibrillar aggregates, mainly appeared in the Aβ1-42 oligomer-rich samples that were prepared according to the protocol (Figure 2). By using the anti-Aβ antibody 6E10, we observed that the number of Aβ1-42 peptides was similar in the HFIP-dissolved Aβ1-42 monomer sample and the Aβ1-42 oligomer-rich sample (Figure 2).
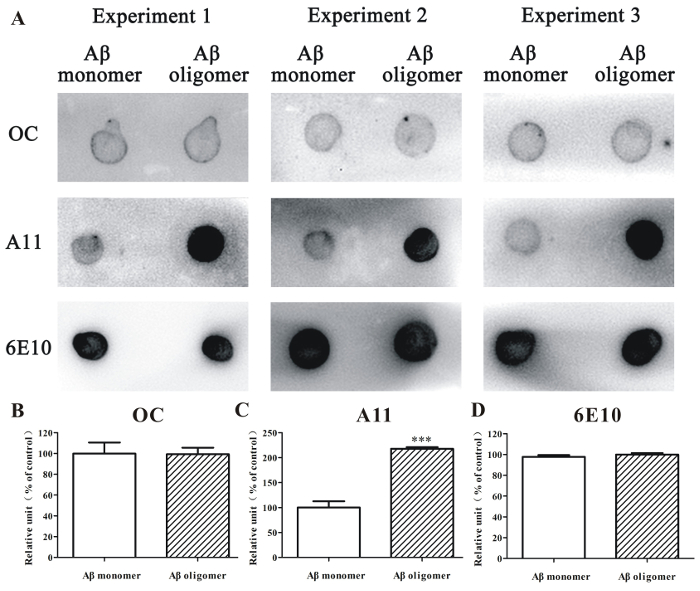
Figure 2. Dot blotting analysis of Aβ1-42 monomer and Aβ1-42 oligomer-rich samples. (A) The HFIP-dissolved Aβ1-42 monomer sample (10 µM) and the Aβ1-42 oligomer-rich sample (10 µM) prepared according to this protocol were examined by dot blotting analysis using the anti-oligomer antibody A11, anti-fibrillar oligomer antibody OC, and anti-Aβ antibody 6E10. (B – D) The semi-quantitative analysis of grayscale was performed by ImageJ. The data, expressed as a percentage of control, were the mean ± SEM of 3 independent experiments; ***p <0.001 vs. the Aβ1-42 monomer group (ANOVA and t-test). Please click here to view a larger version of this figure.
To further investigate if this protocol could be used to screen inhibitors of Aβ1-42 oligomers, curcumin, a known Aβ oligomer inhibitor, was used. We found that a co-incubation with curcumin (0.1 – 1 µM) significantly reduced the relative amounts of A11-positive Aβ1-42 oligomer, as evidenced by the decreased staining of A11 in the curcumin co-incubated Aβ1-42 oligomer sample than in the normal incubated Aβ1-42 oligomer-rich sample (Figure 3). At the same condition, curcumin (0.1 – 1 µM) did not alter the number of Aβ1-42 peptides, as the staining of 6E10 was even in all samples (Figure 3).
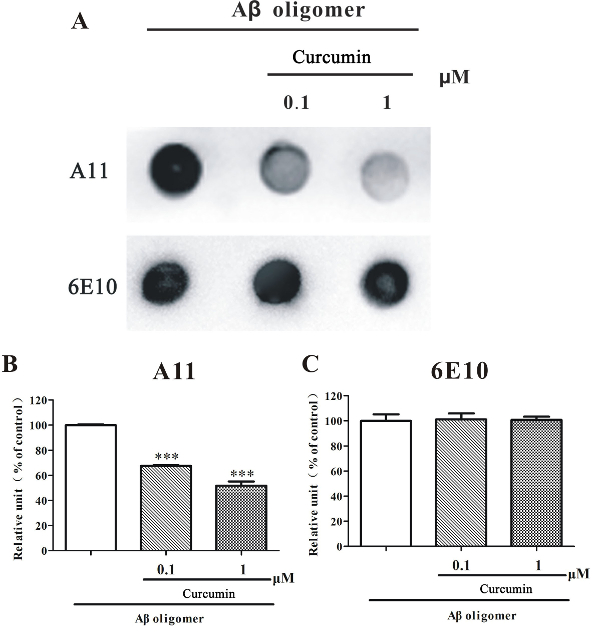
Figure 3. Dot blotting analysis of curcumin co-incubated Aβ1-42 oligomer-rich samples. (A) HFIP-dissolved Aβ1-42 monomers (10 µM) were prepared according to this protocol and incubated with or without curcumin (0, 0.1, 1 µM) under shaking for 48 h. The samples were examined by a dot blotting analysis using A11 and 6E10. (B) Semi-quantitative grayscale analysis was performed by ImageJ. The data, expressed as a percentage of control, were the mean ± SEM of 3 independent experiments; ***p <0.001 vs. the Aβ1-42 alone group (ANOVA and Tukey's test). Please click here to view a larger version of this figure.
To verify oligomer formation, we have also used a non-denaturing gel. We found that oligomer (>4 KD) was present in the Aβ1-42 oligomer-rich sample, while most monomer (4 KD) was found in the Aβ1-42 monomer sample (Figure S1).
We have investigated Aβ aggregates in the supernatant and pellet of the Aβ1-42 sample. After 48 h of incubation, Aβ1-42 oligomer but not Aβ1-42 fibrillar aggregates were found in the supernatant of the sample as determined by A11, OC, and 6E10 antibodies (Figure S2). At the same condition, Aβ1-42 fibrillar aggregates but not Aβ1-42 oligomers were found in the pellet of the sample (Figure S2).
We have also examined the morphology of the curcumin-treated Aβ1-42 sample by using TEM. The curcumin-treated sample contains a few spherical spots, suggesting that curcumin could reduce the amount of Aβ1-42 oligomer (Figure S3).
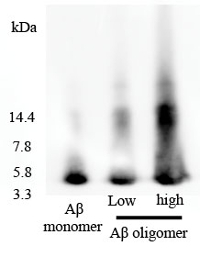
Figure S1. Non-denaturing gel electrophoresis analysis of Aβ1-42 monomer and Aβ1-42 oligomer-rich samples. The HFIP-dissolved Aβ1-42 monomer sample (10 µM) and the Aβ1-42 oligomer-rich sample (5 µM, low; 10 µM, high) prepared according to this protocol were examined by non-denaturing gel electrophoresis using the anti-Aβ antibody 6E10. Please click here to view a larger version of this figure.
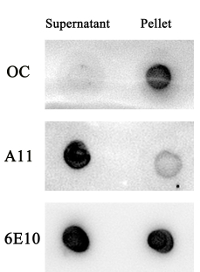
Figure S2. The supernatant of Aβ1-42 solution contains mainly oligomer, but not fibrillar aggregates. An Aβ1-42 solution (50 µL), after shaking for 48 h, was centrifuged at 18,000 x g for 10 min. The supernatant was collected and the pellet was re-dissolved in 850 µL of double-distilled water. The supernatant and pellet were further examined by a dot blot assay using OC, A11, and 6E10 antibodies. Please click here to view a larger version of this figure.
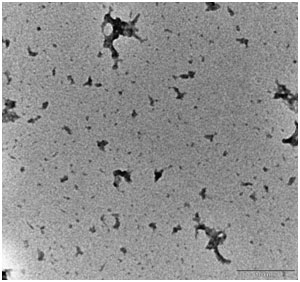
Figure S3. TEM analysis of curcumin co-incubated Aβ1-42 oligomer-rich samples. HFIP-dissolved Aβ1-42 monomers (10 µM) were prepared according to this protocol and incubated with 1 µM curcumin under shaking for 48 h. The samples were examined by TEM. Scale bar = 100 nm. Please click here to view a larger version of this figure.
