The egg-lay chamber was specifically designed to enable P. maidis females to feed while ovipositing in a protective medium from which their eggs could easily be recovered. Using this method, sufficient quantities of precellular embryos were recovered for microinjection with DNA, RNA, and/or proteins. Adult P. maidis females usually lay eggs inside the leaf tissue of the corn plants, which makes getting enough eggs in a short amount of time a challenge because it requires a lot of leaf dissection. The artificial egg-laying environment provides a solution to overcome these problems. As shown in Table 1, 6,483 eggs were collected from a total of 645 females in 4 weeks. Females usually start laying eggs after day 2 and provide most eggs from day 4 to day 6. Oviposition activity slowed down by day 9. Each oviposition chamber was set up on Friday and checked for eggs from Sunday until the next Sunday. Following this schedule allowed most eggs to be collected for microinjections during the work week.
The first practical application of this egg-laying system was to test the efficacy of Cas9-mediated gene knockout, using the P. maidis ortholog of the eye-color gene, white (Pmw), as a target. Mutations in white are known to result in substantial loss of eye pigmentation in other insect species, and White is cell-autonomous, allowing mutations to be detected in injected individuals16,17. To increase the chance that even a small mutation might result in loss of function, guide RNAs were designed to cut within the region of the ATP-binding cassette, which is necessary for White function16. P. maidis embryos were injected with either 20% phenol red (injection buffer), injection buffer with Cas9 at a final concentration of 800 ng/µL (Cas9 control), or Cas9 in injection buffer along with three guide RNAs added at a concentration of 400 ng/µL each. The combination of three guides within one injection mix was intended to further maximize the chances of generating mutants, both by creating a large deletion, and by compensating for the possibility that any one guide might be ineffective for cutting. The developmental rates for each treatment were comparable (Table 2), with 50-60% of injected individuals showing signs of development. Hatch rates for the buffer and Cas9 controls were also comparable; however, the hatch rates of individuals receiving the three-guide mix was relatively lower. At this time, it is unclear if the reduced survival is the result of the loss of white function or the result of unintended consequences of the three-guide mix, such as off-target effects (see the discussion section). However, none of the individuals with complete loss of eye pigmentation (i.e., complete knockout) hatched, and none of the progeny of injected individuals had white eyes. The on-target efficacy of Cas9-based mutagenesis was verified two ways. First, injectees were screened for loss of eye pigmentation.
Of the 71 guide-injected individuals that developed, 23 showed some degree of pigment loss (Figure 10), and 9 of those individuals hatched, resulting in a knockout rate of ≥32%. No eye pigment loss was observed in either control treatment. Second, chromosomal mutations were confirmed via polymerase chain reaction (PCR)18 and sequencing19. Because a mutant line could not be recovered, genomic DNA was analyzed from pools of embryos injected with either the three-guide mix or buffer. The three-guide mix is expected to remove ~180 base pairs from the white locus. This can be seen in the PCR products amplified off of genomic DNA isolated from injected individuals, as well as the associated sequence data generated from those products (Figure 11). This combined evidence indicates that embryos were injected before cellularization occurred.

Figure 1: Aspirator. An effective aspirator can be assembled from attaching a vacuum pump at the intake, via plastic tubing, to a 15 mL plastic conical tube. Approximately 0.5 cm should be carefully removed from the bottom of the conical tube. A cotton ball should be placed in the conical tube, over the opening of the plastic tubing, to catch P. maidis adults as they are collected and keep them out of the vacuum pump. Please click here to view a larger version of this figure.

Figure 2: Construction of adult containers. (A) The supplies needed (clockwise from top left): screen, hot glue gun, razor blade, 1 oz container. (B) A large hole should be cut in the bottom of the 1 oz container, and a square of screen is cut just large enough to cover this hole. (C) The screen is then glued over the hole using hot glue. (D) Once the glue is set, any excess mesh should be removed. Please click here to view a larger version of this figure.

Figure 3: Sexing P. maidis adults. The ventral sides of male (left) and female (right) P. maidis adults are shown. The ovipositor, visible over the female abdomen, is the clearest indicator of the sex of an individual. Please click here to view a larger version of this figure.

Figure 4: Sealing adults into containers. (A) A 5 cm x 5 cm square of plastic paraffin wax film. (B) The film should be stretched evenly to 3-4 times its original size. (C) Once adults have been put into the adult container, the stretched film should be placed over the opening to secure the adults. A 400 µL drop of 10% w/v sucrose solution should then be placed on top of the film. (D) To provide adequate feeding pressure for the adults, a second 5 cm x 5 cm square of plastic paraffin film should be similarly stretched and placed over the drop of sucrose. Please click here to view a larger version of this figure.
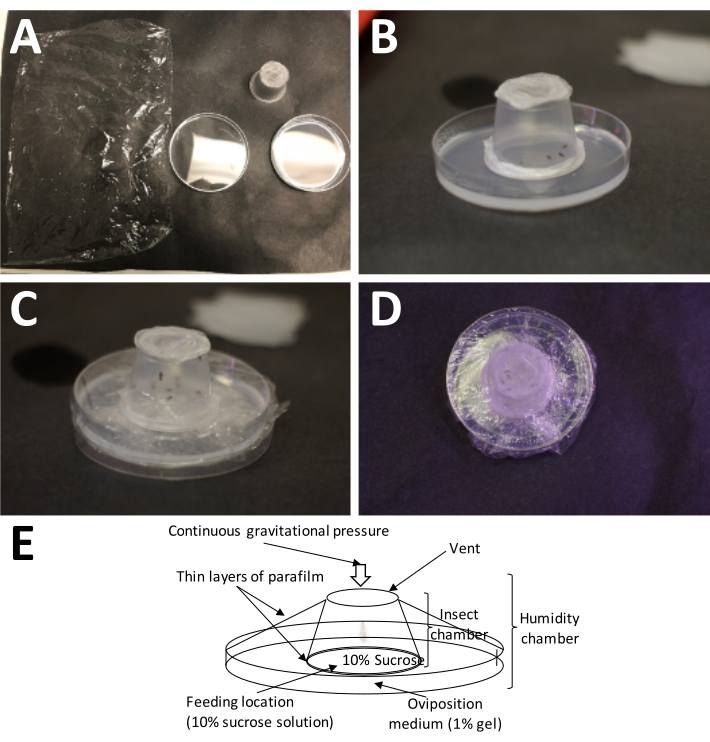
Figure 5: Setting up an oviposition chamber. (A) The supplies needed (clockwise from top left): plastic wrap, a completed adult container (with adults), and a Petri dish with 1% agarose (oviposition medium). (B) The adult container should be placed onto the agarose with the plastic paraffin film/10% sucrose 'sandwich' placed directly on the oviposition medium. (C) Plastic wrap is used to secure the adult container to the oviposition medium. This keeps the medium from drying out too quickly. (D) Care should be taken to avoid covering the screen of the adult container, so that air exchange may still continue. (E) Diagram of the oviposition chamber. Please click here to view a larger version of this figure.

Figure 6: Humidified hood. A hood outfitted with a humidifier has been set up around the injection scope to minimize air drafts and maintain humidity while the embryos are being handled. Flaps can be folded over the entrance after the worker is in place, to aid in maintaining proper humidity levels. Please click here to view a larger version of this figure.
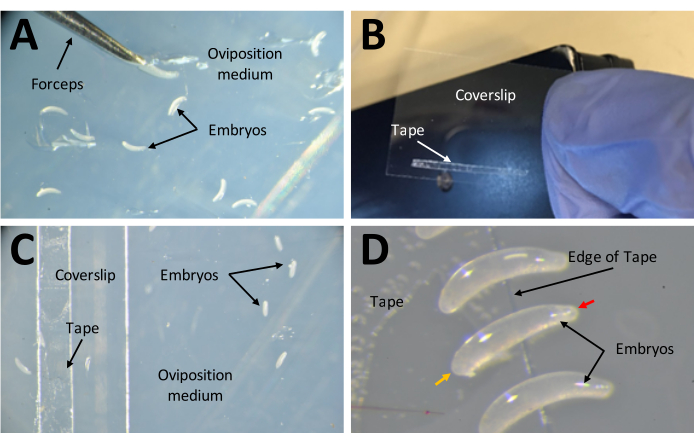
Figure 7: Collecting embryos in preparation for injections. (A) Embryos that have been deposited in the oviposition medium. A pair of fine forceps are used to extract embryos from the medium and place them on its surface. (B) A narrow strip of 1 mm x 15 mm double-sided tape on a 22 mm x 30 mm coverslip. (C) The coverslip can be placed on the oviposition medium for ease of transferring embryos from the surface of the medium to the tape on the coverslip. (D) P. maidis embryos are banana shaped, with one end narrower than the other (narrow end indicated with red arrow head; wider end indicated with yellow arrow head in example embryo). The wide end of the embryo should be placed on the tape. Please click here to view a larger version of this figure.
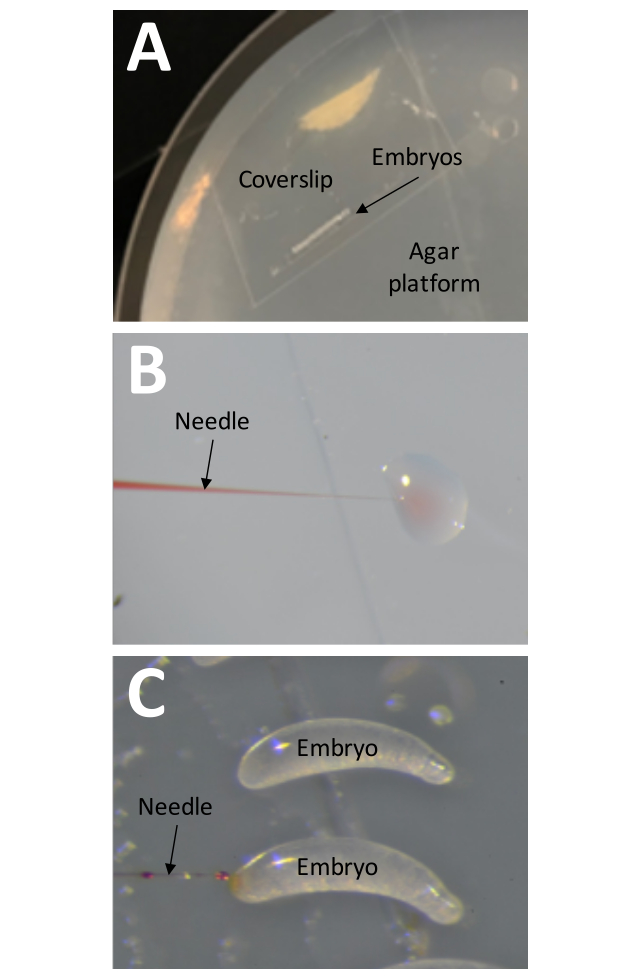
Figure 8: Injection. (A) The injection platform is a Petri dish filled to the brim with 1% agar. The coverslip with a strip of tape holding embryos should be placed directly on the surface of the injection platform. (B) The injection pressure should be tested before embryos are injected by 'injecting' a small amount of injection solution into a drop of water. This method can also be used at any time during the injection process to check the needle for clogs. (C) Embryos should be injected by inserting the needle into the larger end of the embryo. Injection solution should be visible if the injection was successful. Please click here to view a larger version of this figure.

Figure 9: Post-injection care. (A) Once all the embryos on a coverslip have been injected, the coverslip should be placed in a fresh Petri dish containing 1% agarose. (B) The Petri dish with the coverslip can then be maintained in a humidity chamber (like the one shown) until embryos hatch. Please click here to view a larger version of this figure.
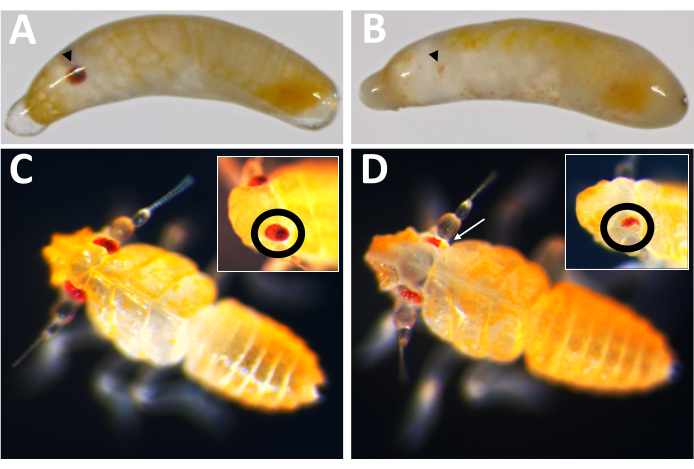
Figure 10: Pmw knockout phenotype. (A) Age-matched control and (B) Pmw knockout embryos, with developing eyes indicated by black arrowheads. The embryo in B is mosaic, as a small stripe of pigmentation can be seen. (C) Age-matched control and (D) Pmw knockout hatchlings, with insets showing a different angle on the eyes. The hatchling in D is also mosaic. A white arrow points to an area in the main picture showing loss of pigmentation. Please click here to view a larger version of this figure.
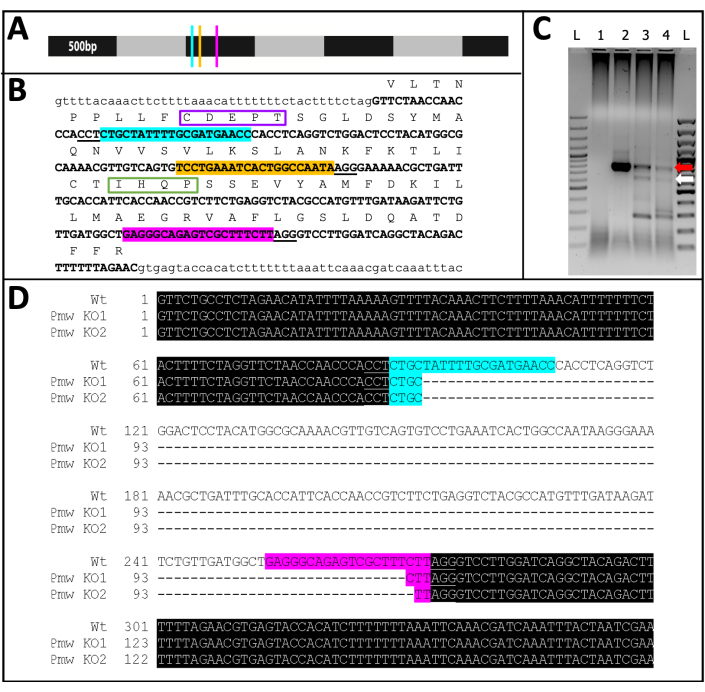
Figure 11: Pmw knockout sequence. (A) To-scale model of Pmw mRNA, marked in 500-bp increments, with locations of gRNA binding sites indicated: G1, blue; G2, yellow; G3, pink. Any frame-shift mutations generated at this point will disrupt the majority of the translation product. (B) Genomic context of gRNA sites, all in one exon (bold capitalized text). Guide binding sites are highlighted in the same colors as A, and PAMs are underlined. Span is ~300 bp. The in-frame translation of the exon is shown above, as single-letter abbreviations in capital text. Two motifs specific to eye pigment transporters are marked. The CDEPT motif of the Walker B functional domain is boxed in purple, and the IHQP motif of the H-loop domain is boxed in green. Both domains are critical to ATP-transporter function. (C) The Pmw target region was amplified using two rounds of PCR. The second-round product was examined on a gel for evidence of size-shift due to successful removal of the region between the guides. Lanes: L = 100 bp ladder; 1 = PCR water control; 2 = Buffer injected eggs; 3-4 = two separate sets of eggs injected with three-guide mix. Only embryos receiving the three-guide mix produced both the WT band (red arrow) and the band resulting from a complete excision (white arrow). (D) To confirm the identity of the lower (white arrow) band, this DNA was purified, cloned, and sequenced. The top line is the wild-type sequence. The other two lines are sequences from two clones. Three additional clones matched the bottom sequence. Blue highlight indicates the binding site of Guide 1, while pink highlight indicates the binding site of Guide 3. In both alleles, the entire region between these two guide sites has been deleted. Abbreviations: Pmw = Peregrinus maidis white gene; gRNA = guide RNA; PAM = protospacer adjacent motif; ATP = adenosine triphosphate; PCR = polymerase chain reaction; WT = wild-type; KO = knockout. Please click here to view a larger version of this figure.
| Set | # of cups | # of females in each cup | # of eggs | Total # of eggs | |||||||
| Day 2 | Day 3 | Day 4 | Day 5 | Day 6 | Day 7 | Day 8 | Day 9 | ||||
| 1 | 10 | 15 | 0 | 26 | 166 | 355 | 530 | 193 | 91 | 37 | 1398 |
| 2 | 15 | 15 | 22 | 238 | 489 | 699 | 520 | 379 | 203 | 58 | 2608 |
| 3 | 8 | 15 | 0 | 57 | 230 | 190 | 116 | 80 | 34 | 1 | 708 |
| 4 | 10 | 15 | 0 | 226 | 446 | 519 | 301 | 179 | 24 | 15 | 1710 |
| Total | 43 | 15 | 23 | 547 | 1331 | 1763 | 1467 | 831 | 352 | 111 | 6483 |
Table 1: Representative egg collections from artificial oviposition environment. Data from four sets of egg-collection cups are shown, with egg tallies starting on the second day after setup and running through the ninth day.
| Injection Treatment | Total Injected | Total Developed | Total Hatched | Development Rate (%) | Hatch Rate (%) |
| Buffer | 39 | 20 | 12 | 51 | 31 |
| Cas9 | 39 | 24 | 14 | 61 | 36 |
| Cas9 +Pmw gRNAs | 121 | 71 | 28 | 59 | 28 |
Table2: Survival and knockout rates from injections of 3 different injection mixes.
