The mediator release assay, based on huRBL cells (Figure 1A and B), results in a bell-shaped dose-response curve (Figure 1C). For simplified data representation, the antigen concentration necessary for the half maximum mediator release can be calculated using linear regression (Figure 1D). A cell viability assay is performed to exclude cytotoxic effects derived from either the sensitizing serum or the antigen used for stimulation (Figure 1E). The assay can be used to test the reactivity of different sera to a certain antigen. In our case, 4 out of 5 sera, derived from birch pollen allergic patients, responded to Bet v 1 stimulation. Serum #1 showed the highest mediator release (Figure 2). Serum #5 did not respond to Bet v 1 stimulation and, thus, might react to other birch pollen allergens (e.g., Bet v 2, profilin). These data indicate that Bet v 1 is a potent allergen responsible for IgE-mediated allergic symptoms. By using the huRBL assay, the cross-reactivity of IgE to homologous allergens can be assessed (Figure 3). Here, both birch pollen allergic patients responded well to Bet v 1, whereas only patient #2 responded also to Cor a 1, the Bet v 1-homologous food allergen found in hazelnuts. Based on these data, patient #2 most likely has higher Cor a 1-cross-reactive IgE levels than patient #1, resulting in oral allergy symptoms upon hazelnut consumption. Even the assessment of the hypoallergenic nature of mutant variants of allergens (decreased potency) can be analyzed and compared to their wild-type counterpart (Figure 4). In the provided example, the release curve of the fold variant shifted towards a higher antigen concentration compared to the wild-type allergen, resulting in a significantly higher concentration of antigen necessary to provoke half maximal release (Figure 4B). These data imply that the generated mutant/fold variant is less allergenic compared to the wild-type protein. This reduced potency to trigger IgE-mediated degranulation highlights the hypoallergenic character of the fold variant. Based on this assay, the fold variant is an interesting candidate for allergen-specific immunotherapy since it might cause reduced IgE-associated side effects during the treatment.
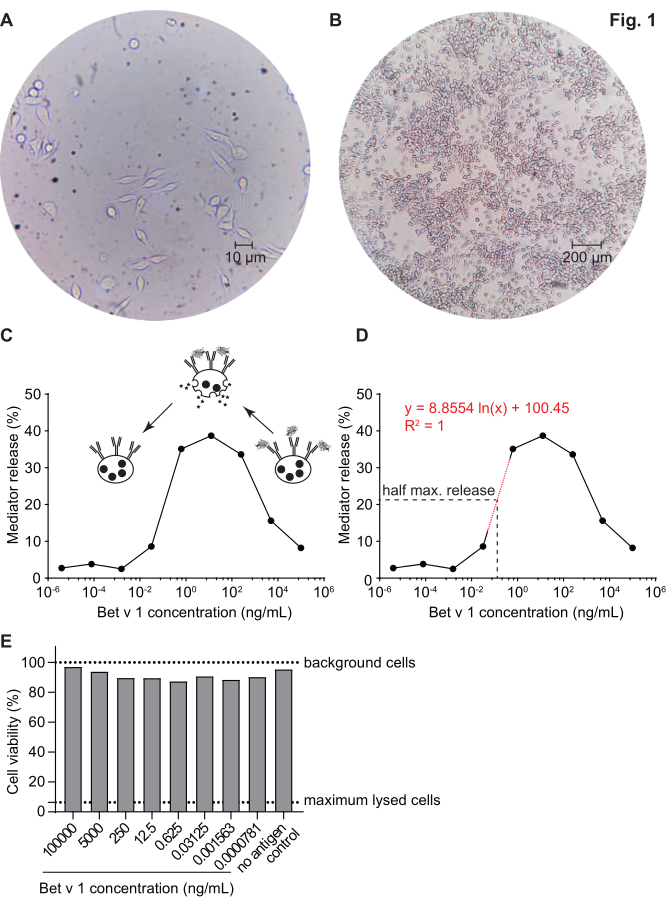
Figure 1: Humanized RBL cells and a representative bell-shaped curve of IgE-allergen cross-linking-induced β-hexosaminidase release. RBL cells are adherent to the culture flasks, which gives them a rod-like shape as they are trying to attach themselves (A). An ideal level of confluence for cells to be harvested is no more than 90% (B). Cells are shown under magnification of 40x and 10x, respectively. Cells that were sensitized with human serum of a birch pollen allergic individual reacting upon challenge with recombinant Bet v 1 (rBet v 1), the major birch pollen allergen (C). As surrogate for mediator release, the β-hexosaminidase activity is measured in cell supernatants. The bell-shaped curve results from a monovalent occupation of antigen epitopes on IgE due to the excess of allergen, which competitively inhibits the allergen-IgE cross-linking at high antigen concentrations. Another explanation for the low release at high allergen concentrations is the inhibition of intracellular pathways in presence of excess antigen. For determination of the allergen concentration necessary to obtain half maximal release, a logarithmic regression line based on the experimental values representing the linear part of the slope of the mediator release curve was used (D). The red dotted line represents the logarithmic regression line used for calculation. The formula of the regression line is shown in red. The half maximal release is defined as: half maximal release = (minimum release value + maximum release value)/2. In the example, the calculated half maximal release was 20.6%. The representative human serum used in this experiment was diluted 1:20 for incubation with huRBL cells, and the antigen concentration used for stimulation ranged from 100 µg/mL to 0.004 pg/mL of Bet v 1. A cell viability assay, in this case a MTT assay, was performed with the remaining cells after antigen stimulation to assess the influence of the sensitizing serum as well as of the antigen dilution on cell viability and cell count (E). Untreated background cells and lysed cells (maximum lysis) are shown as dotted line. Please click here to view a larger version of this figure.
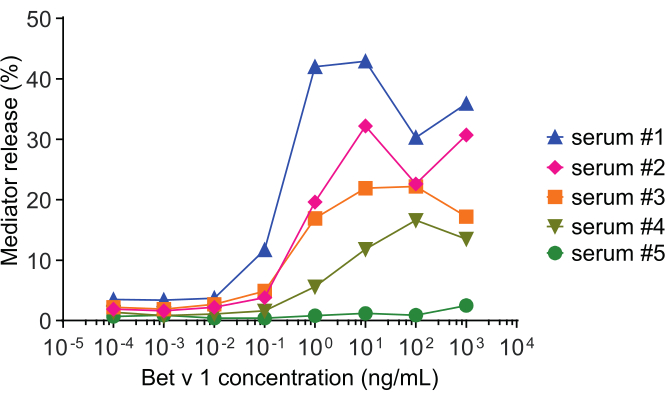
Figure 2: Representative curves of β-hexosaminidase percent release of five different human sera. The same antigen concentration range of rBet v 1 was incubated with huRBL cells that were sensitized with sera of different birch pollen sensitized individuals. There is a clear difference of percent release between the different patients corresponding to the severity of their symptoms. Notice that patient #5 is non-reactive to the major birch pollen allergen Bet v 1. All five human sera used to obtain these mediator release curves were diluted equally 1:20 for incubation with huRBL cells. Please click here to view a larger version of this figure.
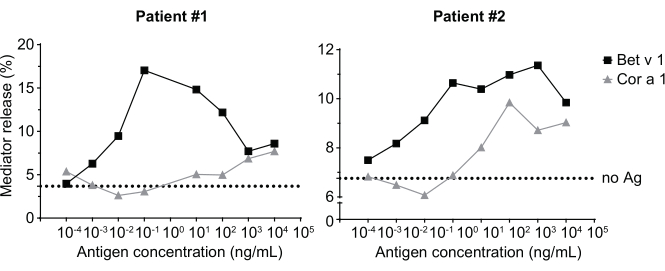
Figure 3: Cross-reactivity of IgE derived from sera of birch pollen sensitized patients with the Bet v 1 homologous hazelnut allergen Cor a 1. Two representative sera of patients sensitized to birch pollen strongly react to Bet v 1 as well as to a lesser degree to the homologous allergen Cor a 1. Patient 2 is shows a significant reaction to Cor a 1, and thus will likely exhibit oral allergy symptoms upon hazelnut consumption, compared to patient 1 where the mediator release is almost negligible. The dotted line represents the no-antigen control, which are cells sensitized with the human sera but not stimulated with an allergen, and, thus, serves as indication for the bottom signal plateau/background. Please click here to view a larger version of this figure.
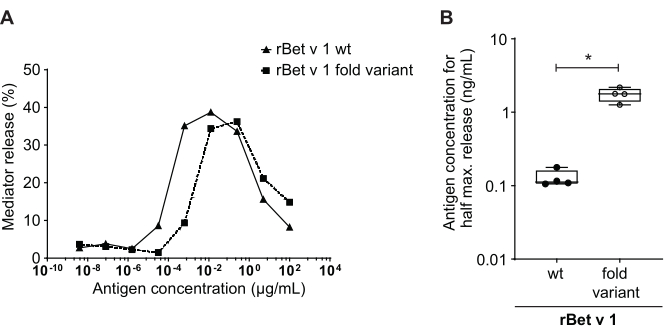
Figure 4: Comparison of percent release between rBet v 1 wild type and a hypoallergenic fold-variant. The same serum of a birch pollen-sensitized individual was incubated with rBet v 1 wild type (wt) and a hypoallergenic fold-variant of the major birch pollen allergen (A). Even though mediator release is seen in both antigens, there is a clear shift toward higher antigen concentrations when comparing the fold-variant to wild type rBet v 1 for the same percent release. A standard way of comparing the difference in percent release of different antigens is calculating the concentration of antigen needed to attain half maximum release (B). This is usually performed in biological replicates (testing of the same antigen range for each allergen in different human sera). Usually, in order to draw any significant conclusions, the mediator release is performed with sera from at least 8 to 10 different patients. Here the results of four different sera are plotted as an example. A paired t-test was used for statistical analysis. *p ≤ 0.05; **p ≤ 0.01; ***p ≤ 0.001; ****p ≤ 0.0001. Please click here to view a larger version of this figure.
| Potential questions and troubleshooting | Solution |
| Assay-to-assay variability due to altered cell responsiveness | Ensure that the cell passage cycle number does not exceed 20 to 30 passages. Make frozen stocks at early passages for future experiments. |
| Rather rely on biological replicates (use of different sera) than technical ones. | |
| Sera contains low levels of specific IgE | A lower final serum dilution can be used (i. e. 1:10) instead of 1:20. Conversely, sera containing high levels of specific IgE can be further diluted (1:30 or 1:40). |
| Not enough cells to perform the assay | Make sure the confluency in a T-75 flask is around 50-90%. Passage more flasks. |
| Cytotoxic effects of sera, i. e. due to incomplete complement-inactivation | Perform a cell viability assay in addition to the mediator release assay. Increase Ag8 concentration to avoid incomplete complement-inactivation. |
| Low signal | Improve signal-to-noise ratio of the assay by diluting the 1x Tyrode´s buffer in deuterium oxide (D2O) instead of dH2O, or by using a sera with higher levels of specific IgE for the allergen of interest. |
| Allergen is not stable in Tyrode’s buffer (e. g. precipitation) | Make stability tests in 1x Tyrode’s buffer prior to the assay procedure. Substitution of Tyrode’s buffer is not recommended. |
| Problems finding the right starting concentration for the respective allergen | Adaption of dilution series to cover the full release curve (more dilution steps, 1:20 dilution instead of 1:10). |
| Poor assay performance indicated by low/no signal | Avoid cytotoxic effects from either sera or antigen stimulation (e.g. enzymatic allergens). Wash and soak the cells carefully. Avoid exposure to air for too long and prevent cells from drying out. |
| How do I know if the bottom signal plateau is reached? | Add "no antigen" controls to your plate. These are sensitized cell, which were only stimulated with 1x Tyrode´s buffer but without an allergen. |
| Do I need a positive control in addition to the maximum lysis wells? | As additional positive control a serum and antigen combination known to cause degranulation can be used or an anti-FcεR1 antibody. |
| How many wells do I need? | That depends on your titration series, the number of antigens and sera you want to analyze. Plan the layout for the 96-well plates according to how many sera/antigens you are going to test. Do not forget to add the "no antigen controls", the background cells (non-sensitized, non-stimulated) as well as the maximum lysis wells. |
| How many sera should I test? And do I need replicates? | Although the assay is quite robust, there is some assay-to-assay variability due to altered cell responsiveness. Therefore, it is recommended to rather rely on biological replicates (using different sera) than on technical replicates. A minimum of eight different sera is sufficient to analyze allergens. However, as shown in Fig. 4B, significant results can already be obtained using less sera. |
Table 1: Troubleshooting.
