rSARS-CoV-2/Nluc infection in K18 hACE2 transgenic mice (Figures 1 and 2)
Figure 1A shows a schematic representation of the rSARS-CoV-2/WT (top) and rSARS-CoV-2/Nluc (bottom) used to assess infections in vivo. Figure 1B shows the schematic flow chart applied to assess rSARS-CoV-2/Nluc infection dynamics in K18 hACE2 transgenic mice. Four-to-six-week-old female K18 hACE2 transgenic mice (N = 4) were either mock-infected with 1x PBS or infected with 105 PFU of rSARS-COV-2/WT or rSARS-CoV-2/Nluc intranasally. At 1-, 2-, 4- and 6-days post-infection, mice were sedated using the isolation chamber and then injected with Nluc substrate retro-orbitally. The isolation chamber was immediately placed in the IVIS and Nluc signal was assessed in vivo using the imaging software. Nluc expression was readily detected in mice infected with rSARS-CoV-2/Nluc but not those infected with rSARS-CoV-2/WT, or mock-infected (Figure 2A). Quantitative analyses showed Nluc intensity at different days post-infection (Figure 2B). Gross lesions on the lung surface of mice infected with rSARS-CoV-2/Nluc were comparable to those in the rSARS-CoV-2/WT infected group (Figures 2C). Lastly, mice organs (lungs, nasal turbinate, and brain) were homogenized, and viral titers were determined by plaque assay (PFU/mL) and Nluc activity was determined using the luciferase assay following the manufacturer's instructions. Plaques were assessed by immunostaining using the cross-reactive SARS-CoV N monoclonal antibody 1C7C7. Viral titers detected in the rSARS-CoV-2/Nluc infected mice were comparable to those infected with rSARS-CoV-2/WT in all organs at different days post-infection (Figure 2D). Nluc activity was only detected in the organs from rSARS-CoV-2/Nluc-infected mice (Figure 2E). A separate group of mock-infected and virus-infected mice were monitored for 12 days for changes in body weight (Figure 2F) and survival (Figure 2G). Mice infected with rSARS-CoV-2/Nluc and rSARS-CoV-2/WT lost up to 25% of their body weight and all succumbed to viral infection between 7-8 days post-infection (Figure 2F–G).
rSARS-CoV-2/Venus infection in K18 hACE2 transgenic mice (Figures 3 and 4)
Figure 3A shows a schematic representation of the rSARS-CoV-2/WT (top) and rSARS-CoV-2/Nluc (bottom) used to assess infections ex vivo. Figure 3B shows the schematic flow chart applied to assess rSARS-CoV-2/Venus dynamics in K18 hACE2 transgenic mice. Four-to-six-week-old female K18 hACE2 transgenic mice (N= 4/group) were either mock-infected with 1x PBS or infected with 105 PFU of rSARS-COV-2/WT or rSARS-CoV-2/Venus intranasally. At 1-, 2-, 4- and 6-days post-infection, mice were euthanized, and their lungs were excised and imaged ex vivo using an IVIS. Venus expression was readily detected in all lungs from mice infected with rSARS-CoV-2/Venus but not those infected with rSARS-CoV-2/WT, or mock-infected (Figure 4A). Quantitative analyses showed that Venus intensity peaks at 2 days post-infection and decreases over the course of infection in the lungs of infected mice (Figure 4B). Images of the lung surface revealed gross lesions of mice infected with rSARS-CoV-2/Venus was comparable to that of rSARS-CoV-2/WT infected mice (Figure 4C). Finally, mice organs (lungs, nasal turbinate, and brain) were homogenized, and viral titers were determined by plaque assay and assessed by immunostaining using the SARS-CoV N protein cross-reactive monoclonal antibody 1C7C7. Infection with rSARS-CoV-2/Venus resulted in comparable viral titers to those observed in mice infected with rSARS-CoV-2/WT in all organs (Figure 4D). A separate group of mock-infected and virus-infected mice were monitored for 12 days for changes in body weight (Figure 4E) and survival (Figure 4F). Mice infected with rSARS-CoV-2/Venus and rSARS-CoV-2/WT lost up to 25% of their body weight and all succumbed to viral infection by day 9 post-infection with no survival (Figures 4E-4F).
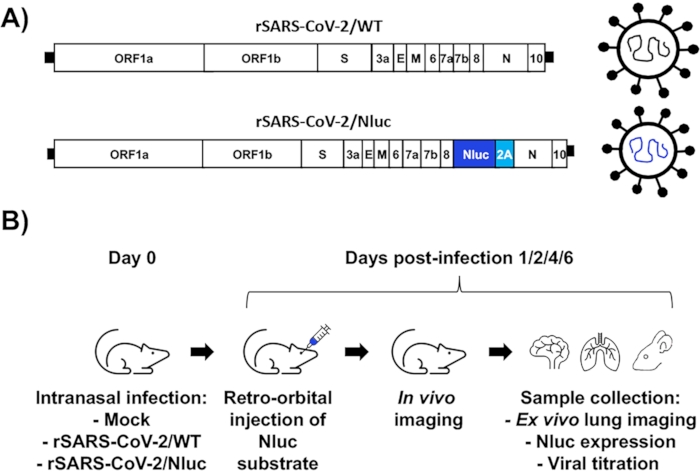
Figure 1: Assessment of rSARS-CoV-2/Nluc infection in vivo using K18 hACE2 transgenic mice. (A) Schematic representation of rSARS-CoV-2/WT (top) and rSARS-CoV-2/Nluc (bottom). (B) Schematic flow chart for the assessment of rSARS-CoV-2/Nluc in vivo. Please click here to view a larger version of this figure.
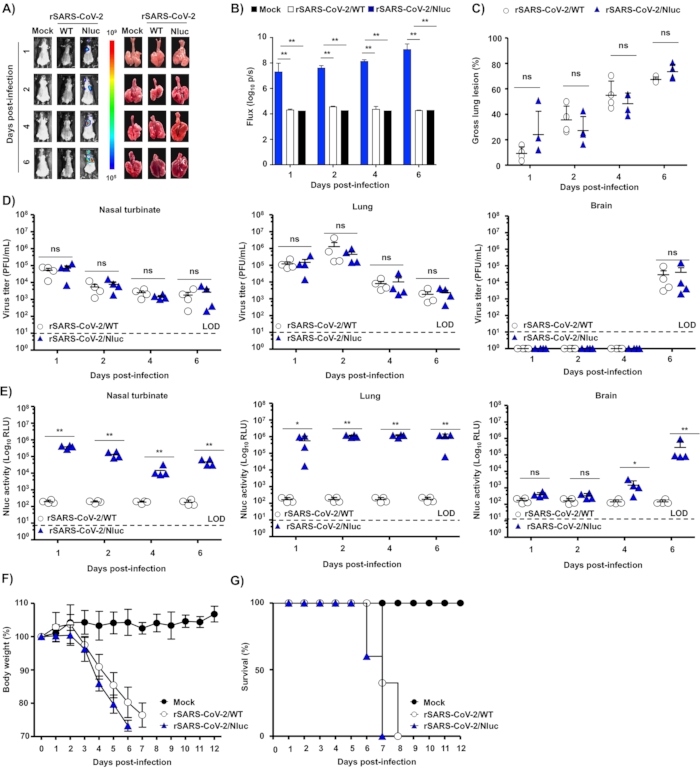
Figure 2: rSARS-CoV-2Nluc expression in infected K18 hACE2 transgenic mice. (A–B) Four-to-six-week-old female K18 hACE2 transgenic mice were mock-infected (N = 4) or infected with rSARS-CoV-2/WT (N = 4) or rSARS-CoV-2/Nluc (N = 4) using 105 PFU per animal. The mice were anesthetized at 1-, 2-, 4- and 6-days post-infection, after being retroorbital injected with the Nluc substrate. (A) Nluc expression was determined under an in vivo imaging system, and lungs from mock-infected and infected mice were excised and photographed at 1-, 2-, 4- and 6-days post-infection . (B) Nluc intensity was quantitively analyzed by the image analysis software and (C) gross lesions on the lung surface were quantitively analyzed by ImageJ (C) **P < 0.01. (D) Viral titers in the nasal turbinate (left), lungs (middle), and brain (right) from mice infected with rSARS-CoV-2/WT and rSARS-CoV-2/Nluc were determined by plaque assay. (E) Nluc activity in the nasal turbinate (left), lungs (middle) and brain (right) were measured under a luciferase multi-plate reader. ns, not significant.Mock- and virus-infected mice were monitored for 12 days for changes in (F) body weight and (G) survival. All data are presented as mean ± SD for each group and analyzed by SPSS13.0 (IBM). A P value of less than 0.05 (P < 0.05) was considered statistically significant. This figure has been modified from Ye C. et al.41. Please click here to view a larger version of this figure.
Figure 3: Assessment of rSARS-CoV-2/Venus infection in vivo using K18 hACE2 transgenic mice. (A) Schematic representation of rSARS-CoV-2/WT (top) and rSARS-CoV-2/Venus (bottom). (B) Schematic flow chart for the assessment of rSARS-CoV-2/Venus in vivo. Please click here to view a larger version of this figure.
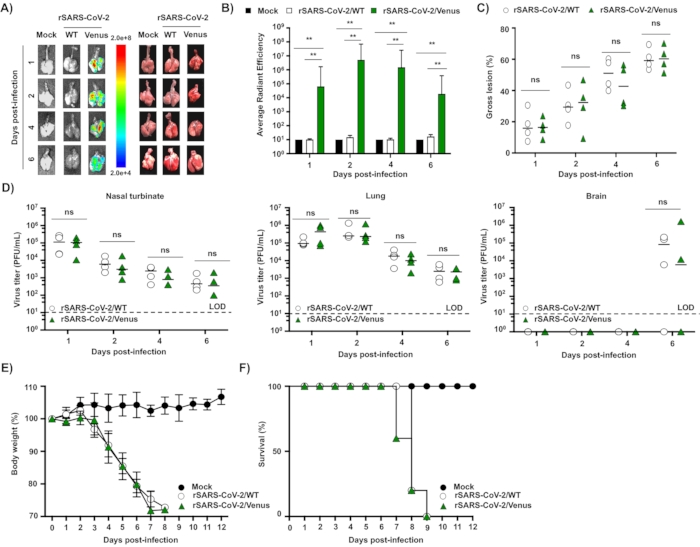
Figure 4: rSARS-CoV-2/Venus expression in infected K18 hACE2 transgenic mice. (A–B) Four-to-six-week-old female K18 hACE2 transgenic micewere mock-infected (N = 4) or infected (105 PFU/mouse) with rSARS-CoV-2/WT (N = 4) or rSARS-CoV-2/Venus (N = 4). Lungs were excised at 1-, 2-, 4-, and 6- days post-infection, images of lungs were photographed at 1-, 2-, 4- and 6-days post-infection. (A) Venus expression was assessed under an IVIS, (B) fluorescence intensity was quantitively analyzed by the image analysis software and (C) the gross lesions on the lung surfaces were quantitively analyzed by ImageJ. **P < 0.01. (D) Viral titers in the nasal turbinate (left), lungs (middle) and brain (right) from mice infected with rSARS-CoV-2/WT and rSARS-CoV-2/Venus were determined by plaque assay. ns, not significant. Mock- and SARS-CoV-2-infected mice were monitored for 12 days for (E) body weight loss and (F) survival. All data are presented as mean ± SD for each group and analyzed by SPSS13.0 (IBM). A P value of less than 0.05 (P < 0.05) was considered statistically significant. This figure has been modified from Ye C. et al.41. Please click here to view a larger version of this figure.
