1. Source of fungal strains
- Use South African isolated fungal strains of both M. pinghaense 5 HEID (GenBank Accession number: MT367414/MT895630) and M. robertsii 6EIKEN (MT378171/MT380849), collected from apple orchards in the Western Cape province, South Africa.
- Grow cultures of each EPF isolate on 60 g of Sabouraud dextrose agar medium, supplemented with 1 g of yeast extract (SDAY) and 10 µL of Streptomycin.
NOTE: Incubate EPF cultures at a controlled temperature of ± 25 °C in the dark.
2. Metarhizium pinghaense and M. robertsii conidial suspension inoculation
- Preparation of the solid substrates
- Use two agricultural products, namely flaked oats and flaked barley, as growth mediums for the two EPF isolates and autoclave bags (305 mm × 660 mm) as fermentation bags.
- Use a small polyvinylchloride waste pipe (1000 mm × 50 mm) to create a neck for the fermentation bag at the open end of the autoclave bag and use autoclave tape to secure the pipe to the bag.
- Close the pipe with a sterile cotton wool plug to allow sufficient gas exchange during fermentation.
- Weigh dry grains (200 g) of both the flaked oats and flaked barley and place them in the fermentation bags, and to each bag, add 100 mL of distilled water and thoroughly mix the contents of the bags.
- Leave the wet grains to rest for 15-30 min to absorb moisture before autoclaving and sterilization. Prepare six bags for each grain type, and to prevent contamination, place the bags in other autoclave bags, and autoclave the substrates at 121 °C for 55 min.
- Preparation of conidial suspension inoculum
- Harvest 2-3-week-old fungal conidia from fungal cultures of both M. pinghaense and M. robertsii by scraping, using a sterile surgical blade.
- Suspend the collected fungal conidia in 20 mL of sterile distilled water, supplemented with 0.05% Tween 20, and vortex-mix the conidial suspensions for 2 min.
- Prepare 20 mL of conidial suspensions, with a concentration of 1 × 107 conidia/mL, and inoculate the flaked oat and flaked barley solid substrates, respectively.
NOTE: Use a hemocytometer to determine conidial concentrations.
- Inoculation of the solid substrates
- Open each bag by removing the cotton wool neck plugs, and add the prepared 20 mL conidial suspension to the cooled autoclaved substrate.
- Close the bags again, using the neck plugs, and massage the contents of the bag to allow the fungal inoculum to become evenly mixed with the grain substrate.
- Incubate the fermentation bags at a controlled temperature of ± 25 °C, and ensure sufficient gaseous exchange between the culture and the environment.
NOTE: This procedure must be conducted under a laminar flow cabinet.
- Fermentation phase
- Manually massage the grain substrates in the fermentation bags 2 days after the inoculation and incubation, when visible mycelial growth occurs and the substrate has begun to be clumped by the growing fungus.
NOTE: This is done to mix the inoculated granules to allow for mycelial branching to take place during the first early stages of the vegetative growth of the fungi. - Use a kitchen rolling pin to manipulate the substrate bed to avoid the physical heterogeneity of the grain substrate beds and the bed thickness of the substrate mass.
NOTE: The process promotes fungal metabolism, which optimizes fungal spore production, and maximizes the surface area, promoting conidial yield18. - Leave the fermentation process to continue for up to 4-5 weeks, and check the fermentation bags every 2 days for the presence of any white vegetative overgrowth that can develop during the fermentation process, which can greatly affect the fungal conidia yield.
NOTE: Immediately terminate fermentation in the fermentation bags containing any white vegetative overgrowth, and dry the fungal cultures18.
- Manually massage the grain substrates in the fermentation bags 2 days after the inoculation and incubation, when visible mycelial growth occurs and the substrate has begun to be clumped by the growing fungus.
3. Blastospore inoculation
- Blastospore production and inoculation
- Prepare a liquid culture medium, containing 1 L of distilled water, 30 g of glucose, 20 g of yeast extract, 4 g of potassium phosphate diabasic (K2HPO4), 15 mL of corn steep liquor, and 10 µg/mL of the antibiotic Streptomycin, for both the M. pinghaense and the M. robertsii.
- First, heat the distilled water, switch off before reaching boiling point, and add each of the ingredients, except for Streptomycin, to the hot water in the pot. Bring the medium to a gentle boil for 3-4 min, and constantly stir the medium to allow for the proper mixing of the ingredients and prevent the settling of some of the ingredients at the bottom of the pot.
- Pour a total of 100 mL of the medium into nine different 250-mL flasks and place a cotton wool plug on each flask, and cover the cotton wool with aluminum foil as a stopper (Figure 1A).
- Autoclave the medium in the flasks for 55 min at 121 °C. Following the autoclaving, allow the medium in the flasks to cool and add 10 mg/mL of Streptomycin to the medium in each flask (Figure 1B).
- Collect two to three bacterial loops of fungal conidia from 2-3-week-old fungal culture plates for both the EPF isolates, M. pinghaense and M. robertsii, and transfer to each 100 mL liquid media in the 250-mL flasks, under sterile conditions, and seal the flasks.
- Incubate the liquid culture flasks at ± 25 °C, on an orbital shaker at 140 rpm for 3 days, and cease the incubation once the cultures show signs of high turbidity with fungal blastospore growth (Figure 1C).
- To detect any possible bacterial contamination from the cultures, draw a 100 µL sample from each liquid culture flask after 24 h during the incubation and plate on three SDA plates per isolate. Incubate the plates for 48 h, at a controlled temperature of ± 25 °C.
- Preparation of the solid substrate
- Use parboiled long-grain white rice as a solid substrate medium for the blastospores of both M. pinghaense and M. robertsii (adapted from Jaronski and Jackson18).
- Prepare the fermentation bags as detailed above, in steps 2.1.1-2.1.3, and for each bag, add 1 kg of rice and 300 mL of sterile distilled water.
- Gently mix the contents of the fermentation bags and place in outer autoclave bags in an upright position, and autoclave at 121 °C for 55 min. Following the autoclaving, allow the substrates to cool down for ± 45 min under sterile conditions.
- Inoculation and fermentation
- Remove the closure of each of the liquid culture flasks of both M. pinghaense and M. robertsii under a laminar flow and flame the rim of each flask for 10 s.
- Pour the 100-mL liquid cultures into the fermentation bags by removing the cotton wool plugs from their necks (Figure 2). Place the cotton plugs back on and cover the top of the bag's neck with surgical paper secured with a rubber band.
NOTE: Determine blastospore concentration for each flask using a hemocytometer, and use blastospore concentrations of 1 × 107 – 5 × 108 blastospores/mL to innoculate the substrates28. - Twist the top part of the bag and mix the contents of the bag by shaking and light manipulation of the substrate by massage, and incubate the bags at ± 25 °C, by flattening the substrate in the bags to prevent the formation of thick beds18.
- Break the substrate in the fermentation bags by massaging contents of the bags, 2-3 days following the inoculation and the incubation, when visible mycelial growth and the binding of the substrate by the fungus had occurred (adapted from the technique of Jaronski and Jackson18).
NOTE: Allow fermentation to continue for 4 weeks.

Figure 1: Liquid culture medium in 250-mL flasks. (A) Before autoclaving. (B) After autoclaving and inoculation with EPF spores. (C) Turbid medium with fungal blastospores. Please click here to view a larger version of this figure.

Figure 2: Prepared blastospore liquid culture medium. (A) Metarhizium robertsii and (B) Metarhizium pinghaense prior to the inoculation of rice as a solid substrate. Please click here to view a larger version of this figure.
4. Drying of fungal cultures
- Dry the fungal cultures for 10-12 days following fermentation, prior to their use in trials, by transferring the sporulated cultures into 26-30 kg (30 x 43 x 15250 cm3) brown paper bags.
- To improve the tensile strength of the paper bags, horizontally cut off one-third of the top part of each bag, and line the bottom of the bag (Figure 3A, B).

Figure 3: Preparation of paper bags, drying procedure of cultures, and packaging. (A,B) The preparation of brown paper bags. The drying procedure of Metarhizium species cultures grown on (C,E) parboiled rice and (D,F) flaked barley. (G) Paper bags closed with staples to create a triangular tent structure. Please click here to view a larger version of this figure.
- Gently crumble the substrate in each fermentation bag, cut off the corner of each bag and transfer the whole culture to the paper bags through the space left by the cut-off corner (Figure 3C-F). To avoid the excessive escaping of fungal spores into the air, perform this process slowly.
NOTE: Conduct this process under sterile conditions, using laminar flow, to avoid contamination. - Label each paper bag and fold over the top end of each bag twice and close with staples to create a triangular tent structure, and place the bags on wire drying racks to allow proper, even drying, at a controlled temperature of ± 25 °C and low humidity of 30-40%.
- Turn the bags daily to allow even drying of the cultures and to avoid any vegetative regrowth that might take place, which would lower the yield of harvestable fungal spores.
- Weigh each drying bag after every 2 days during the drying process, and continue the drying process for each bag until little to no change is observed in the mass of the bags between the successive days.
5. Harvest of fungal conidia
- Harvest fungal conidia mechanically from the cultures using three nested sieves, a test sieve (ETS) mesh no. 35 (with 500-µm aperture), nested on a test sieve (with 212-µm aperture), nested on ETS mesh no. 100 (with 150-µm aperture), mounted on a collection pan.
- Load the dry culture sample on the ETS mesh no. 35 sieve slowly and place a lid on the sieve to prevent the release of fungal conidia into the air.
- Add 10-12 glass marbles to the sieves to assist the passage of the fungal conidia through the mesh screens and avoid the retention of the conidia in the sieve, which can result in reduced spore recovery.
- Tape and seal the sieve joints using electrical tape to prevent the escape of conidial dust.
- Place the sieves on a vibratory shaker, fitted with a sticky pad, to secure the collection pan and sieves, for 20-25 min, at a motion of 560-640 vibrations per min (Figure 4).
NOTE: Technique adapted from Jaronski and Jackson18.

Figure 4: Harvesting of fungal spores from dried Metarhizium robertsii cultures on rice and flaked barley. (A) 10-12 glass marbles added to the sieves to assist the passage of the fungal conidia through the mesh screens. M. robertsii conidia harvested from cultures on (B) rice, and (C) flaked barely. (D) Sieves on a vibratory shaker. Please click here to view a larger version of this figure.
- Remove the test sieves from the collection pan, collect conidia and store the collected conidia in airtight and water-impermeable zipper-lock bags for long-term storage (3-6 months).
6. Quantification of fungal conidia produced
- Measure and record the mass of each grain substrate prior to the harvesting of the fungal conidia from each substrate.
- Measure the overall weight of the collected conidia by subtracting the mass of the collected spores from the mass of the solid substrate.
NOTE: The total conidia yield does involve not only the harvested conidia but also the fungal conidia left on the solid substrate. - Weigh the substrate and remove 10 g from the weighed substrate. Suspend the 10 g of the substrate in 0.05% Tween 20 and dilute in 10 mL of sterile distilled water.
- Vortex-mix the suspension for 2 min, and use a hemocytometer to do the spore count to determine the number of conidia washed from the substrate.
- Conduct further dilutions by transferring 1000 µL of the 10 mL washed conidial suspension to 9 mL of sterile distilled water to make up 10 mL of dilution suspensions.
- Vortex-mix the conidial suspensions for 2 min, and determine the conidial concentrations.
NOTE: Follow the procedure and formula described by Inglis, Enkerli, and Goettel30 to determine the conidial concentration of the suspensions. - Collect and suspend a total of 0.1 g of the collected conidial powder from each culture in 10 mL of sterile distilled water supplemented with 0.05% Tween 20, and vortex-mix the conidial suspension for 2 min, and determine the conidial concentration using a hemocytometer.
- Conduct further dilutions by transferring 1000 µL of the 10 mL conidial suspension to 9 mL of sterile distilled water to make up the 10 mL suspension dilutions.
- Vortex-mix the conidial suspensions for 2 min, calculate the conidial concentrations and determine the number of conidia per gram.
- Multiply the number of conidia per gram of harvested powder by the initial mass of the harvested conidial powder. Multiply the number of conidia washed from the substrate by the total weight of the spent substrate, being the substrate from which the conidia were harvested.
- Add the two given values together and divide by the initial dry weight of the substrate to calculate the number of conidia per kg or g of the substrate18.
NOTE: The calculations were done mainly for the rice substrate. The germination or conidial viability test was conducted for both the M. pinghaense and the M. robertsii isolates to determine the viability of the produced conidia.
7. Data analysis
- Use an appropriate computer software program to conduct the statistical analysis of the obtained results.
NOTE: Statistical analysis of the data was done using STATISTICA version 13.5.0.17.
A decline in the content mass of the cultures on rice for both the M. pinghaense and the M. robertsii was observed over time during the drying stage of the fungal cultures, with no, or little, change being observed in the mass once the cultures were dry (Figure 5). The harvested dry fungal conidia powder of both the M. pinghaense and the M. robertsii is shown in Figure 6.
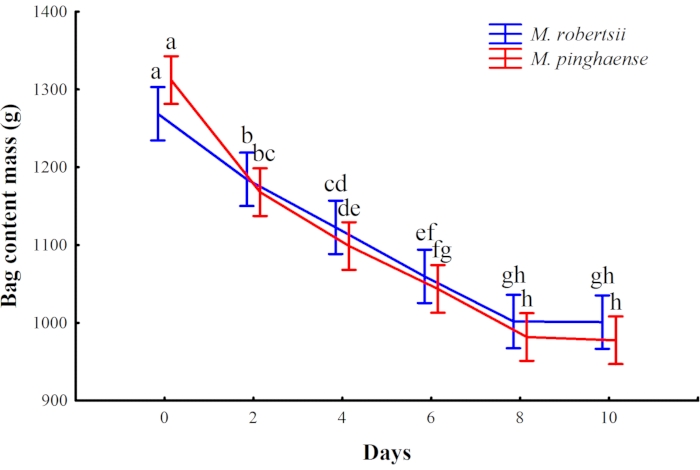
Figure 5: The change in the bag content mass of the Metarhizium robertsii and the M. pinghaense cultures on rice (95% confidence interval) over 10 days (ANOVA; F5,35 = 2.21; p = 0.08). Different letters above the bars indicate the significant difference (p < 0.05) obtained in bag content mass over the days. Please click here to view a larger version of this figure.

Figure 6: Harvested conidia. (A) Harvested conidia in the collection pan. (B) Metarhizium robertsii. (C) M. pinghaense. Please click here to view a larger version of this figure.
No significant difference was observed in the average conidial powder that was harvested from each grain type for both M. pinghaense and M. robertsii. Harvested dry conidia for both had an average of >90% conidial viability for both M. pinghaense and M. robertsii. On the rice substrate, M. pinghaense produced an average of 8.2 g ± 4.38 g of conidial powder, which slightly exceeded the amount that was obtained for M. robertsii (6 g ± 2 g). An average of 1.83 g ± 1.47 g of dry M. robertsii conidia was harvested from the flaked barley substrate, whereas zero M. pinghaense was harvested from the substrate. No dry fungal conidia were harvested from the flaked oats substrate for either M. pinghaense or M. robertsii (Figure 7).
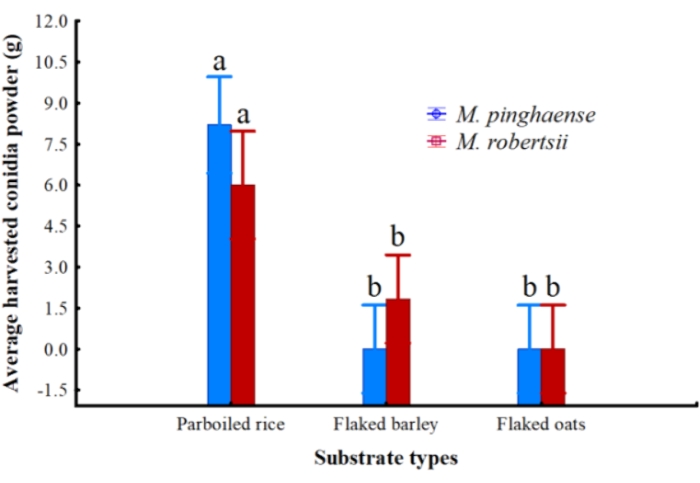
Figure 7: Average amount of conidia powder. The average amount of conidia powder of Metarhizium pinghaense and M. robertsii, measured in grams (95% confidence interval) harvested from the three solid substrates: parboiled rice, flaked barley, and flaked oats (ANOVA: F2,27 = 2.82; p = 0.08). Please click here to view a larger version of this figure.
A significant difference in the estimated average conidia per gram, harvested from the rice grains, was observed between M. pinghaense (3.51 x 107/g ± 0.43 x 107/g) and M. robertsii (4.31 x 107/g ± 0.38 x 107/g). M. robertsii had a slightly higher average conidia count per gram than M. pinghaense (Figure 8).
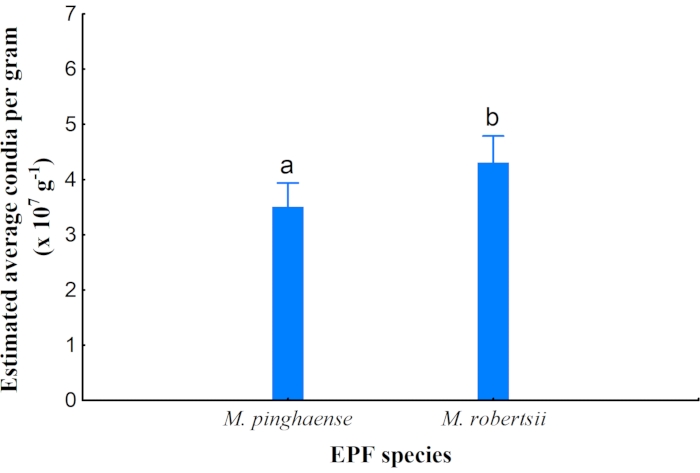
Figure 8: Average conidia per gram. The estimated average conidia per gram for Metarhizium pinghaense and M. robertsii (95% confidence interval) from the rice cultures (ANOVA: F 1,7 = 8.47; p = 0.02). Please click here to view a larger version of this figure.
No significant difference in the estimated number of conidia present on the spent rice substrate was observed between M. pinghaense (5.02 x 107/mL ± 1.47 x 107/mL) and M. robertsii (3.70 x 107/mL ± 0.91 x 107/mL) (Figure 9). However, M. pinghaense had a slightly higher number of conidia present on the rice substrate than M. robertsii.
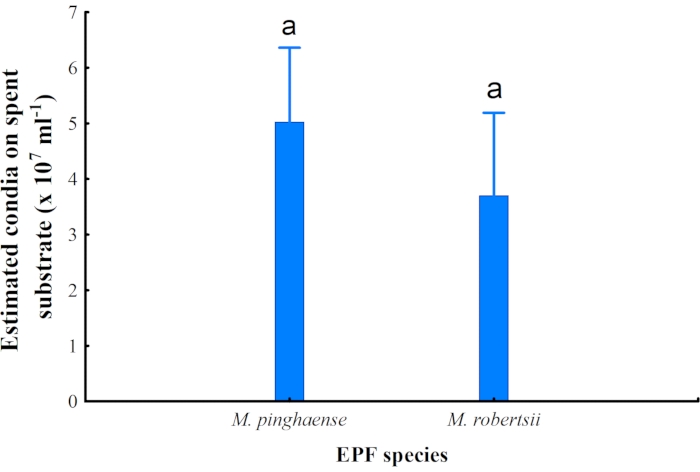
Figure 9: Average fungal conidia. The estimated average fungal conidia of Metarhizium pinghaense and M robertsii (95% confidence interval) from the cultures (F1,7 = 2.45; p = 0.16) present on the spent rice culture substrates. Please click here to view a larger version of this figure.
No significant difference in the estimated overall conidia yield was observed between the M. pinghaense (5.09 x 107/g ± 1.35 x 107/g) and the M. robertsii (3.55 x 107/g ± 0.85 x 107/g) obtained from the rice substrate cultures. However, M. pinghaense produced a slightly higher conidial yield than did M. robertsii, which produced a lower conidial yield (Figure 10).
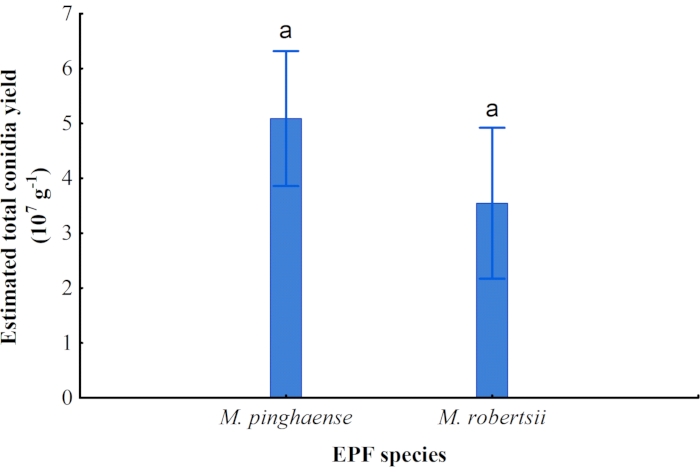
Figure 10: Total conidia yield. The estimated total conidia yield of Metarhizium pinghaense and M. robertsii (95% confidence interval) from the cultures on rice (F1,7 = 3.91; p = 0.09). Please click here to view a larger version of this figure.
