We successfully differentiated human BD-PSCs into endoderm/hepatic progenitor cells and hepatocytes by applying a two-step protocol. Morphological changes during the hepatic differentiation process are shown in Figure 1. BD-PSCs differentiate into hepatocytes going through three different stages. The first stage represents the differentiation into endodermal cells L4, the second, differentiation to hepatic progenitor cells (hepatoblast) L8, exhibiting a typical polygonal morphology, and the third, the maturation to hepatocytes L15-L24.
Immunofluorescence analysis was performed to confirm the hepatic differentiation of BD-PSCs as presented in Figure 2. Strong expression of endoderm/human liver progenitor marker, like alpha-fetoprotein (AFP), a major plasma protein in fetal serum whose concentration is very low in adult organisms and is therefore considered as a marker for hepatocytes' precursor15 and transthyretin (TTR), a major thyroid-hormone binding protein involved in transporting thyroxine from the bloodstream to the brain16 are found in the cells at the first stage of the hepatic differentiation process at L4 to L8. However, their expression decreases at L15, while the expression of albumin (ALB), the most abundant plasma protein produced mainly by the liver and utterly critical for hepatic differentiation, as well as hepatocyte nuclear factor 4 alpha (HNF-4α), a hepatocytes transcription factor that is involved in the expression of liver-specific genes17 appears firstly at L4, rises throughout the differentiation time L4-L15 reaching a strong and stable expression during the maturation time L15-L24.
Cytokeratin 18 (CK18) is a cytoskeletal protein, one of the major components of intermediate filament expressed in the liver18. The results show that, as expected, CK18 expression correlates with mature hepatocytes (L15-L24), and it is not expressed in hepatocyte progenitor cells.
The well-defined protocol for hepatocyte differentiation in 2D cultures enables the engineering of hepatic 3D cultures starting with BD-PSCs.
We demonstrate here that spontaneous aggregation of these cells in low attachment plates containing hepatocyte induction/maturation medium initiates spheroid formation. The growth track was followed by imaging cells at L2, L4, and L7. (Figure 3A) There is a consistent correlation between spheroid volume and the variable number of cells, as presented in Figure 3B.
The liver is an organ in which most of the drugs in the human body get metabolized. Cytochrome P450 is a superfamily of enzymes (monooxygenases) that are of pivotal importance in the processes of drug and cellular metabolism, detoxification of xenobiotics, and homeostasis. To assess the potential functional activity of BD-PSCs derived hepatic spheroids, we analyzed the expression of drug-metabolizing enzymes like CYP3A4 and CYP2E1, members of CYP3 and CYP2 families19.
Most of the drugs that are used today including codeine, cyclosporin A, erythromycin, acetaminophen, and diazepam as well as many steroids and carcinogens, are metabolized due to the activity of CY3A4 enzyme20. CYP2E1 is involved in the metabolism of endogenous substrates like ethylene glycol, benzene, carbon tetrachloride, and particularly the most important highly mutagenic compound like nitrosamine21.
The spheroids that are formed and differentiated according to the protocol at D14, live stained with antibodies to these two enzymes, reveal the potential hepatic functional activity of BD-PSCs derived spheroids (Figure 4).
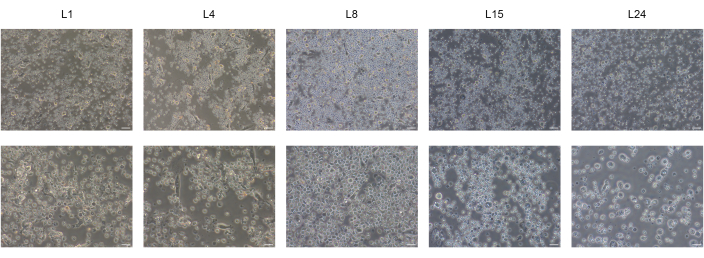
Figure 1: Differentiation of BD-PSCs to hepatic-like cells. Representative micrographs of morphological changes throughout hepatic differentiation of BD-PSCs showing endodermal L4, or polygonal shape L8 morphology finally reaching maturation state at L15 to L24. Scale bars: upper row 50 µm, lower row 20 µm. Please click here to view a larger version of this figure.
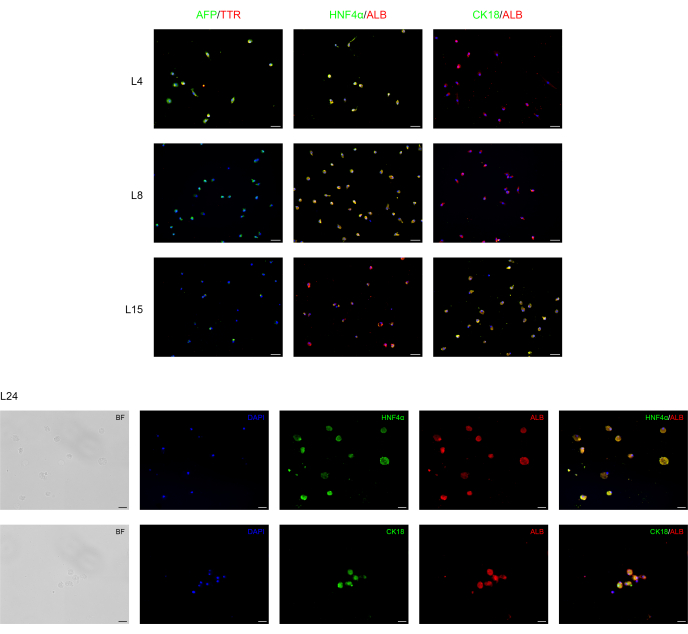
Figure 2: Immunofluorescence analysis of BD-PSCs re-differentiation towards hepatic cells. Endoderm/hepatocytes progenitor and hepatocytes specific markers are expressed during liver differentiation of BD-PSCs in 2D cultures. On days L4 to L8, micrographs show decreased expression of endoderm/hepatic progenitor AFP and TTR while their expression disappeared from L8-L24. Expression of hepatocytes ALB and HNFα markers arise at L4 and increase during maturation, whereas the expression of CK18 appeared first at L15, reaching the maximum at L24. Scale bars for graphs L4-L15: 50 µm and for L24: 20 µm. Control is presented in Supplementary Figure 1. Please click here to view a larger version of this figure.
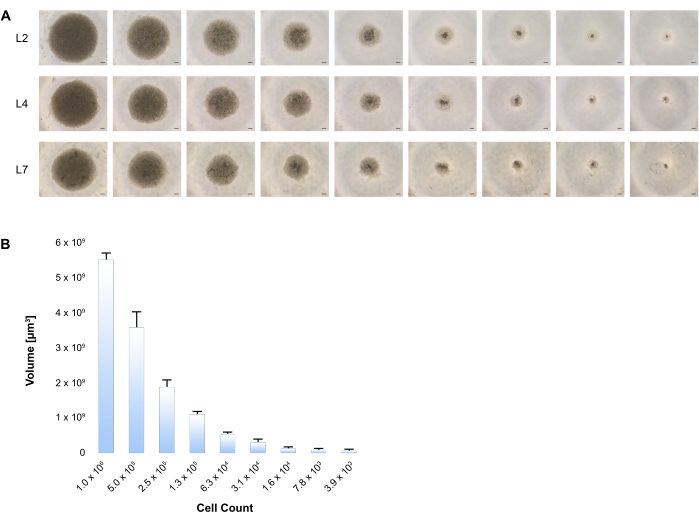
Figure 3: Formation of 3D spheroids upon hepatic differentiation of BD-PSCs. (A) Variable cell numbers of BD-PSCs starting with 1 x 106 to 4000 cells were seeded into low attachment plates, and differentiation was performed according to the two-stage procedure as stated in the Protocol. The generation of 3D human liver spheroids was imaged at different time points, shown are representative phase contrast images at each time during the culture time period. Scale bar: 200 µm. (B) Diameters of at least 4 hepatic spheroids for each size were measured at L4 using microscope imaging software and volumes were calculated. Error bars show standard deviation. Please click here to view a larger version of this figure.
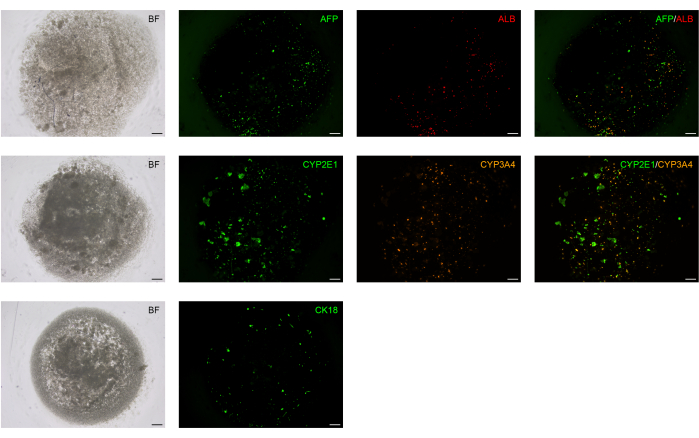
Figure 4: Hepatocyte functional markers are expressed in BD-PSCs-derived liver spheroids. BD-PSCs were differentiated into hepatocytes. Direct immunofluorescence analysis was performed on live cells at L14 using antibodies to ALB, AFP, CK18, and CYP2E1 and CYP3A4, members of the cytochrome P450 family. Scale bar: 200 µm. Please click here to view a larger version of this figure.
Supplementary Figure 1: Negative control for immunofluorescence analysis of BD-PSCs re-differentiation towards hepatic cells. Endoderm/hepatocytes progenitor and hepatocytes specific markers are expressed during liver differentiation of BD-PSCs in 2D cultures. Scale bar: 100 µm. Please click here to download this File.
