All figures in this report were obtained with 201B7-PAX3-GFP iPSCs, in which EGFP replaces one allele of the PAX3 coding sequence in exon 1. Establishment of 201B7-PAX3-GFP iPSCs will be described elsewhere (H. Sakurai, personal communication). The statistical significance was evaluated using statistical software. P-values lower than 0.05 were considered significant.
Characterization of human iPSC-derived PSM and SM cells
To assess the differentiation of human iPSCs toward SM through the PSM state (Figure 2A), FACS analysis, ICC analysis, and RT-qPCR analysis were performed. As shown in Figure 2B, over 85% of the cells were positive for DLL1, a marker of PSM, but negative for PAX3, a marker of SM, after 4 days of PSM induction with human iPSCs. Subsequently, this population became PAX3 positive SM cells after 4 days of SM induction. The PSM-SM transition was also confirmed by ICC (Figure 2C) and RT-qPCR (Figure 2D). TBX6, MSGN1, and WNT3A, PSM markers were expressed at the PSM state (day 4), but not expressed at the SM state (day 8). PARAXIS, MEOX1, and PAX3, SM markers, were expressed at SM, but not expressed at PSM. Furthermore, staining of CDH11, a marker of epithelialized SM, only accumulated at the cell-cell junction, following the addition of SB431542 with CHIR99021 (Figure 2E).
Characterization of SM derivatives induced from human iPSC-derived SM cells
To evaluate the differentiation potency of human iPSC-derived SM, differentiation toward DM, MYO, D, SCL, and SYN (Figure 3A) was assessed by ICC analysis and PAX3 (GFP)-fluorescence. As shown in Figure 3B, DM differentiation was confirmed by ALX4 and EN1 staining, and PAX3 (GFP)-fluorescence; MYO differentiation was confirmed by MYOD, MYOG, and Myosin heavy chain (MHC) staining; D differentiation was confirmed by EN1 and PDGFRa staining; SCL differentiation was confirmed by PAX1, PAX9, and NKX3.2 staining; and SYN differentiation was confirmed by SCX, MKX, COL1A1, and COL1A2 staining.
Characterization of induced D and SYN
1. Enzyme-linked immunosorbent assay (ELISA) for functional analysis of iPSC-derived D
In the human body, one of the primary functions of dermal fibroblasts is to secrete extracellular matrix (ECM) proteins, such as collagen and hyaluronic acid that hydrate the skin and help sustain the skin structure. To demonstrate that a comparable amount of collagen-type 1 and hyaluronic acid proteins were secreted in the culture medium of iPSC-derived D and HDF, ELISA was performed, as shown in Figure 4A.
2. Mechanical stretch stimulation assay for functional analysis of iPSC-derived SYN
As several studies have already reported, mechanical stimulation affects tendon development before and after birth, and promotes the differentiation of tenocytes from precursor cells18,19. Therefore, it is well known that reactivity to mechanical stress is one of the characteristics of tenocytes. To demonstrate the comparable reactivity of human iPSC-derived SYN and human adult tenocytes, a mechanical stretch stimulation assay was performed as shown in Figure 4B.
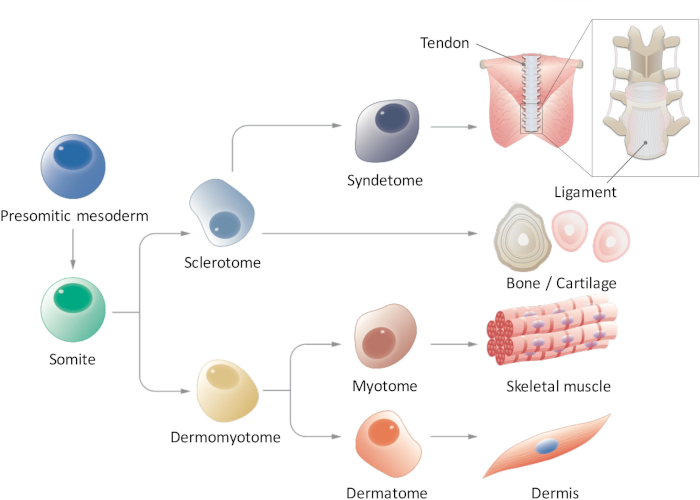
Figure 1: Schematic view of hierarchical differentiation of paraxial mesoderm. Presomitic mesoderm is a cell population that transiently emerges during early embryogenesis and undergoes segmentation to form somites. Somites are a transient stem cell population that gives rise to multiple cell types, such as sclerotome, dermomyotome, syndetome, dermatome, and myotome cells, which eventually differentiate into tendon/Ligament, bone/cartilage, skeletal muscle, and dermis cells. Please click here to view a larger version of this figure.
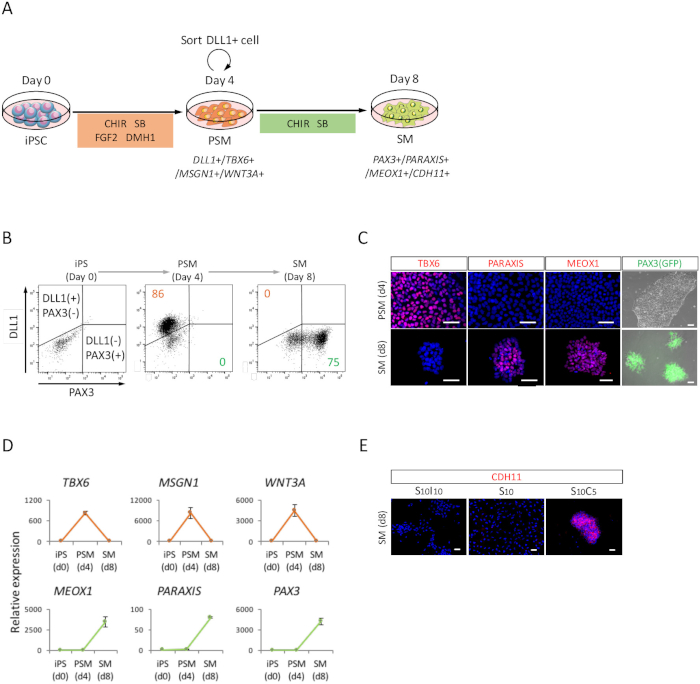
Figure 2: FACS, RT-qPCR, and ICC analysis of human iPSC-derived PSM and SM. (A) Schematic view of a protocol for SM differentiation through PSM. (B) Representative dot plot of DLL1 staining and PAX3 (GFP)-fluorescence on day 4 of PSM induction and day 4 (day 8 from iPSC) of SM induction. (C) Representative immunocytochemical images and PAX3 (GFP)-fluorescence on day 4 of PSM induction and day 4 (day 8 from iPSC) of SM induction. Cells were stained with anti-TBX6, PARAXIS, and MEOX1 antibodies (red) and co-stained with DAPI (blue) or detected with PAX3 (GFP)-fluorescence (green). (D) RT-qPCR analysis of markers for PSM and SM at iPSC, PSM and SM. The means ± standard error (S.E.) from three sets of experiments are shown. (E) Representative immunocytochemical images on day 4 (day 8 from iPSC) of SM, cultured in S10I10 (combination of SB431542 and IWR1, an inhibitor of WNT signaling), S10 (SB431542), and S10C5 (combination of SB431542 and CHIR99021) conditions. Cells were stained with anti-CDH11 antibody (red) and co-stained with DAPI (blue). iPS, induced pluripotent stem cell; PSM, presomitic mesoderm; SM, somite; S10, SB431542 10 μM; C5, CHIR99021 5 μM; I10, IWR1 10 μM; Scale bars = 50 μm. This figure has been modified from Nakajima et al. (2018)15. Please click here to view a larger version of this figure.
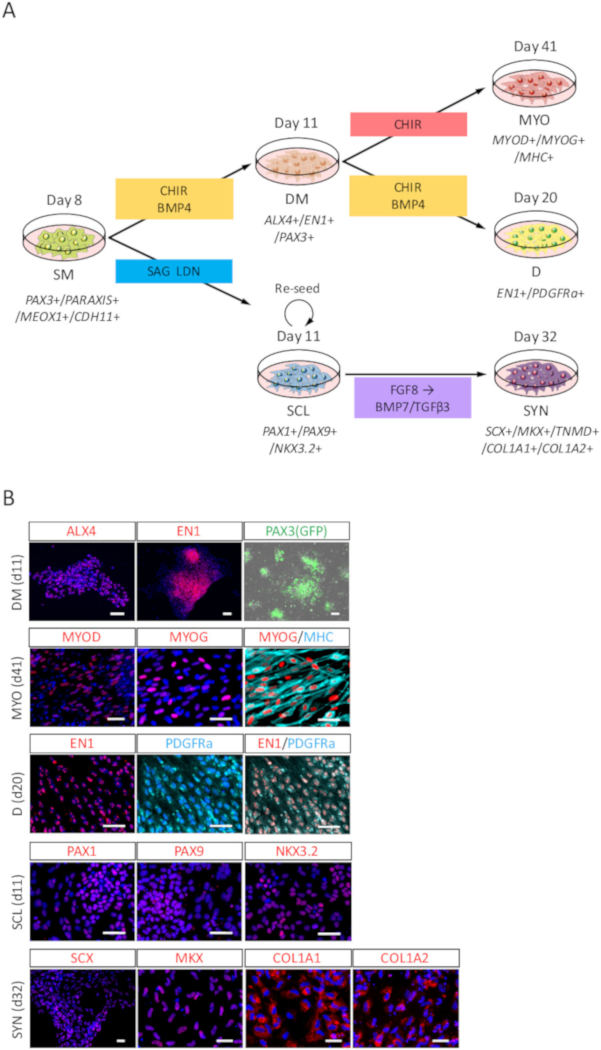
Figure 3: ICC analysis of DM, MYO, D, SCL, and SYN differentiated from human iPSC-derived SM. (A) Schematic view of protocols for SM derivatives differentiation. (B) Representative immunocytochemical images and PAX3 (GFP)-fluorescence on day 3 (day 11 from iPSC) of DM induction, day 30 (day 41 from iPSC) of MYO induction, day 9 (day 20 from iPSC) of D induction, day 3 (day 11 from iPSC) of SCL induction, and day 21 (day 32 from iPSC) of SYN induction. DM, cells were stained with anti-ALX4 and EN1 antibodies (red) and co-stained with DAPI (blue) or detected with PAX3 (GFP)-fluorescence (green); MYO, cells were stained with anti-MYOD, MYOG (red), and MHC (cyan) antibodies, also co-stained with DAPI (blue); D, cells were stained with anti-EN1 (red) and PDGFRa (cyan) antibodies and co-stained with DAPI (blue); SCL, cells were stained with anti-PAX1, PAX9, and NKX3.2 (red) antibodies, and co-stained with DAPI (blue); SYN, cells were stained with anti-SCX, MKX, COL1A1, and COL1A2 (red) antibodies, and co-stained with DAPI (blue). DM, dermomyotome; MYO, myotome; D, dermatome; SCL, sclerotome; SYN, syndetome; Scale bars = 50 μm. This figure has been modified from Nakajima et al. (2018)15. Please click here to view a larger version of this figure.
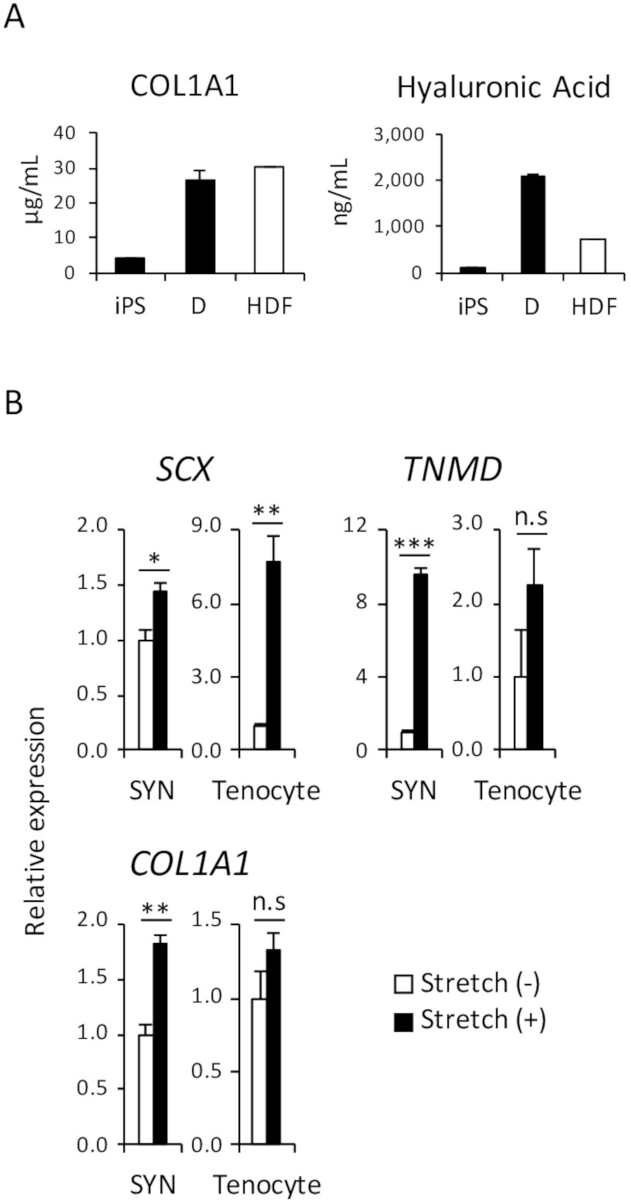
Figure 4: Functional assay of induced D and SYN. (A) The amount of collagen-type 1 and hyaluronic acid proteins in the culture medium were analyzed by ELISA. (B) The effect of mechanical stretch stimulation on induced SYN and human adult tenocytes was assessed by RT-qPCR. The means ± standard error (S.E.) from three sets of experiments are shown. *p < 0.05; **p < 0.01; *** p < 0.001 by Dunnett’s multiple comparisons t-test compared to Stretch (-); n.s, not significant, HDF, human adult dermal fibroblast. This figure has been modified from Nakajima et al. (2018)15. Please click here to view a larger version of this figure.
| Medium/solution | Reagant | Concentration |
| CDM basal medium | Iscove’s modified Dulbecco’s medium/Ham’s F12 | 1:1 |
| Penicillin/Streptomycin | 0.5 % | |
| Chemically defined lipid concentrate | 1 % | |
| Apo-transferrin | 15 mg/mL | |
| Monothioglycerol | 450 mM | |
| Bovine serum albumin | 5 mg/mL | |
| Insulin | 7 mg/mL | |
| CTK solution | Water | – |
| Trypsin | 0.25 % | |
| Collagenase IV | 0.1 mg/mL | |
| Calcium chloride | 1 mM | |
| Knockout SR | 20 % | |
| D induction medium | CDM basal medium | – |
| CHIR99021 | 5 µM | |
| BMP4 | 10 ng/mL | |
| DM induction medium | CDM basal medium | – |
| CHIR99021 | 5 µM | |
| BMP4 | 10 ng/mL | |
| ECM solution | Artificial extracellular matrix | 0.3 mg/mL |
| DMEM/F12 | – | |
| FACS buffer | PBS | – |
| Bovine serum albumin | 0.1 % | |
| Feeder-free cell culture medium | mTeSR1 | – |
| Penicillin/Streptomycin | 0.5 % | |
| HDF culture medium | DMEM | – |
| Fetal bovine serum | 10 % | |
| hESC medium | Primate ES cell medium | – |
| Penicillin/Streptomycin | 0.5 % | |
| FGF2 | 4 ng/mL | |
| MYO induction medium | CDM basal medium | – |
| CHIR99021 | 5 µM | |
| PSM induction medium | CDM basal medium | – |
| SB431542 | 10 µM | |
| CHIR99021 | 10 µM | |
| DMH1 | 2 µM | |
| FGF2 | 20 ng/mL | |
| SCL induction medium | CDM basal medium | – |
| SAG | 100 nM | |
| LDN193189 | 0.6 µM | |
| SM induction medium | CDM basal medium | – |
| SB431542 | 10 µM | |
| CHIR99021 | 5 µM | |
| SYN induction medium-1 | CDM basal medium | – |
| FGF8 | 20 ng/mL | |
| SYN induction medium-2 | CDM basal medium | – |
| BMP7 | 10 ng/mL | |
| TGFβ3 | 10 ng/mL |
Table 1: Media and solution recipes.
| NAME | Forward | Reverse |
| ACTB | CACCATTGGCAATGAGCGGTTC | AGGTCTTTGCGGATGTCCACGT |
| COL1A1 | GGACACAGAGGTTTCAGTGGT | GCACCATCATTTCCACGAGC |
| MEOX1 | GAGATTGCGGTAAACCTGGA | GAACTTGGAGAGGCTGTGGA |
| MSGN1 | GGAGAAGCTCAGGATGAGGA | GTCTGTGAGTTCCCCGATGT |
| PARAXIS | TCCTGGAGAGCTGTGAGGAT | CACACCCTGTCACCAACAGT |
| PAX3 | AGGAAGGAGGCAGAGGAAAG | CAGCTGTTCTGCTGTGAAGG |
| SCX | CCCAAACAGATCTGCACCTTC | GCGAATCGCTGTCTTTCTGTC |
| TBX6 | AGCCTGTGTCTTTCCATCGT | AGGCTGTCACGGAGATGAAT |
| TNMD | CCCTTCATGCTGAAGCCACTT | CTCACTTTCAGCAGAATTGGGG |
| WNT3A | CAAGATTGGCATCCAGGAGT | ATGAGCGTGTCACTGCAAAG |
Table 2: Primer sequences for RT-qPCR analysis.
| Concentration | ||
| 1st Antibody |
ALX4_Goat | 1/50 |
| CDH11_Mouse | 1/1000 | |
| COL1A1_Rabbit | 1/100 | |
| COL2A1_Mouse | 1-2 μg/mL | |
| EN1_Rabbit | 1/50 | |
| MEOX1_Rabbit | 1/50 | |
| MHC_Rabbit | 1/200 | |
| MKX_Rabbit | 1/50 | |
| MYOD_Rabbit | 1/500 | |
| MYOG_Mouse | 1/400 | |
| NKX3.2_Rabbit | 1/50 | |
| PARAXIS_Rabbit | 1/50 | |
| PAX1_Rabbit | 1/50 | |
| PAX9_Rabbit | 1/50 | |
| PDGFRa_Goat | 1/100 | |
| SCX_Rabbit | 1/50 | |
| TBX6_Goat | 1/50 | |
| 2nd Antibody |
Donkey anti Goat IgG(H+L) secondary antibody555 | 1/500 |
| Donkey anti Goat IgG(H+L) secondary antibody647 | 1/500 | |
| Goat anti Mouse IgG(H+L) secondary antibody555 | 1/500 | |
| Goat anti Rabbit IgG(H+L) secondary antibody555 | 1/500 | |
| Goat anti Rabbit IgG(H+L) secondary antibody647 | 1/500 |
Table 3: First and second antibodies for ICC.
