The ministat array described above and in (Figure 1A, B) was used to culture a haploid MATa laboratory strain of budding yeast (S288c) under sulfate-limiting conditions as described previously.10 We tested efficacy for common chemostat applications including determination of physiology and experimental evolution. To validate the ministats, we repeated several experiments previously performed in industrial fermentors modified for chemostat use.10,20,21 ATR Sixfors fermentors were run at a 300 ml working volume, over ten times the volume of the ministats, and have considerably different modes of culture aeration and agitation. We attempted to replicate equipment stability, steady state physiology, experimental evolution results, and gene expression patterns obtained with these fermentors.
Since uniformity of dilution rate and aeration are important aspects of the chemostat design, we measured the actual dilution rate across 32 ministats after 15 generations of growth and found that with a target dilution rate of 0.17 vol/hr (4-5 drops/min) we achieved an average dilution rate of 0.17208 with a standard deviation of 0.0075 across 32 replicates. This range was within our typical tolerance of +/-0.01 vol/hr difference from the target setting, beyond which large-scale changes in gene expression have been observed.22-23 Across 4 ministats the air flow rate was determined to be 307.5 ml/min with a standard deviation of 9.57 ml/min. This suggests that air-flow into the chambers is robust and evenly divided between the 4 chambers and is a value similar to that described for aeration in industrial fermentors.20
We previously observed recurrent amplification of the high-affinity sulfur-ion transporter SUL1 in 8/8 sulfate-limited evolution experiments in yeast.10 Given the consistency of results under this condition we chose sulfate-limitation to test our system’s ability. A requisite element of chemostat culture is the need to maintain a constant chemical environment. Fluctuations in the abundance of a limiting nutrient or other changes in the environment typically result in a change in the number of cells in a given culture. We measured optical density as a proxy for cell number (Figure 2A) and measured reproducibility across 16 replicate cultures, finding an average A600 of 1.12 (~109 cells) after ~15 generations of growth with a standard deviation of 0.057 units, or 5.1%. For comparison, measurements taken from 4 replicate cultures grown in the industrial fermentors showed a standard deviation of 2.5%. Cells were well-mixed in the growth chamber: measurements of OD and cell count taken from the effluent track were equivalent to samples taken directly from the culture tube (data not shown). These results demonstrate the robustness of our platform and ability to maintain a constant chemical environment within a similar tolerance as the industrial fermentor.
As a more sensitive readout of physiology, we compared genome-wide steady state gene expression from cultures grown under sulfate limitation in the ministats and in the industrial fermentor. Gene expression in the ministats showed a high degree of similarity across three biological replicates (Figure 2B). We previously noted that for RNA derived from two replicate Sixfors chemostats and co-hybridized to a microarray, expression of 99% of genes fell within a 1.5-fold range, allowing the use of 1.5X as an empirical significance cutoff.21 Gene expression from three replicate ministats, compared pairwise, showed 99% of genes fell within a 1.5-1.7 fold range, comparable to results from the industrial fermentors. The three samples were hybridized to individual arrays and the pairwise ratios calculated afterwards, so these values include inter-array noise in addition to biological noise, potentially overestimating the variation between replicates as compared to the published, co-hybridized results. 138 genes were differentially expressed >1.5-fold in all three ministat replicates as compared to a sample collected from a matched culture grown in the industrial fermentor. Genes decreased in expression in the ministats were heavily enriched for iron metabolism. This signature may reflect the different metal composition of each device configuration: the Sixfors apparatus includes a metal impeller and aeration assembly immersed in the culture, and the media carboys previously used also required metal hardware. The ministat utilizes stainless steel needles, but no other metal components. Genes with increased expression were largely associated with cell membranes, though the biological significance of this association is unclear.
Finally, we tested experimental evolution under these conditions. After 250 generations of sulfate-limited growth, 4/4 clones tested from 4 independent evolving populations showed amplification of SUL1 as detected by array Comparative Genomic Hybridization (CGH, Figure 2C). This result is consistent with findings in larger volume chemostats over similar time intervals.10

Figure 1. A. Design and arrangement of the ministat array. B. Design of the culture chamber.
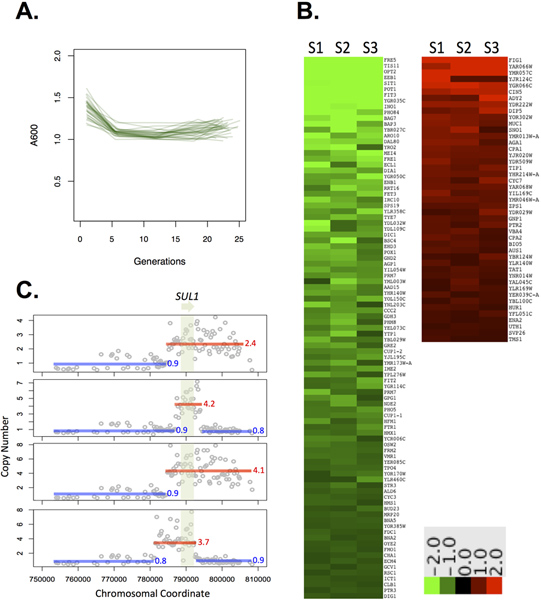
Figure 2. A. Experimental data showing that cultures reach equilibrium within ten generations of growth (n=16). B. Expression data for three biological replicates S1-3 sampled during steady state under sulfate limitation compared to a common reference grown in a matched sulfate-limited Sixfors chemostat. C. SUL1 amplifications recovered in ministats after 250 generations of growth in a sulfate-limited environment. Genomic DNA from each evolved clone was compared to ancestral DNA by CGH as described.21 Mean copy number was calculated for each amplified region and is shown next to each amplicon. All microarray data are deposited in the GEO database under accession GSE36691. Click here to view larger figure.
