The phage demonstrated lytic activity against all tested strains of S. aureus, including MRSA strains, as indicated by the phage spot test. Plaque sizes generally ranged from 5 to 15 mm. No activity was found against other test-cultures (Table 1).
A normal growth of S. aureus ATCC 12600 in NZY medium on shaker-incubator at 37 °C is shown in Figure 1A (a curve labeled by empty circles). The number of bacteria increased from 3.2 x 106 to 4.0 x 108 CFU/ml. Figure 1A (a curve labeled by filled circles) shows results of the co-cultivation of 2.0 x 106 PFU/ml free phage simultaneously added with the same concentration of S. aureus ATCC 12600. Phage, immobilized on the gold surface, demonstrated the lytic activity comparable with the activity of phage in suspension (Figure 1A, curve labeled by top down triangles). Immobilized phage remained infective when the gold piece with phage was first used in 24-hr growing experiment, then it was washed 5 times and stored in PBS for 6 days at 4 °C, and finally reused with a fresh bacterial suspension. (Figure 1A, curve labeled by top up triangles). It seems that immobilized phages inject their DNA in the bacteria, leaving void capsids, which are incapable to infect new bacteria. We hypothesize that immobilized phages on a gold surface served as primary “catalyzers” to infect a few bacteria, which generate free phages those in turn infect new bacteria and so on. Therefore, the most of immobilized phages were probably not used in a first 24 hr growing experiment and, therefore, can be utilized a second time. The surface density of phages deposited by physical adsorption was about ~0.7 phage particle/μm2. This concentration is high, but it can grow up to 10 times 2. Therefore, the gold plate could incorporate some of the free viable phages released by infected bacteria in the first 24 hr growing experiment.
Figure 1B shows the lysis zone around the gold piece, indicating that immobilized phages were capable of lysing bacterial cells. In the control experiments, the empty gold plate did not inhibit bacterial growth. Hence the effective decrease of bacterial growth found at the co-culture of bacteria and immobilized phage is a result of primary interaction of water suspended bacteria and bound phage as shown in Figure 1C. The phage detachment from the gold surface during co-incubation of bound phage and bacteria was small. In order to estimate how many phages were detached from the gold surface during incubation, samples with bound phage were immersed in NZY solution, shaken in the shaker-incubator at 37 °C for 8 hr, and a free phage concentration in supernatant was assessed. We determined that 4.1 x 107 PFU were bound to a 60 mm2 gold piece (~0.7 phage particle/μm2), when only 3 x 104 PFU were detached (0.007% detachment).
The transmission and scanning electron micrographs of intact lytic phage 12600 on the gold substrate surface is shown in Figures 2A and 2B. When the phage suspension was subjected to chloroform treatment, the phage physical appearance was changed. The tail contracted in length and thickened. The polygonal head became rounded (Figures 2D and 2E). We termed the structurally modified lytic phage “spheroid” similar to the name of the chloroform treated filamentous phage 26. In spite of the significant structural changes resulted from chloroform treatment, the spheroid lytic activity measured by plaque numbers has not changed (Figures 2C and 2F). The mean lytic activity of phage and spheroids were (7.4±1.5(SD)) x 1010 (N=4) and (7.5±1.0(SD)) x 1010 (N=7), PFU/ml. At the 0.05 level, the activities of phage and spheroids were not significantly different (Figure 2C and 2F).
The QCM sensors with immobilized lytic phages showed no significant changes in the resonance frequency or energy dissipation when they were exposed to MRSA (Figure 3A). These data indicate that MRSA/phage interaction resulted in a no mass change according to the QCM, and yet the electron micrographs of post assayed biosensors revealed significant bacterial binding at the sensor surface (Figure 3B).
When MRSA suspensions were injected into the flow cell with phage spheroid biosensors, a substantial decrease in the frequency (Δf ≈ -105 Hz) and an increase in the dissipation (ΔD ≈ 26 DU) were observed (Figure 3C). Following phage spheroid-bacterial (MRSA) binding interactions, assayed sensors were exposed to PBP2a antibody conjugated latex beads suspensions. These MRSA assayed biosensors responded to the PBP2a antibody conjugated latex beads suspensions, since a further decrease in the frequency (Δf ≈ -45 Hz) and an increase in the dissipation (ΔD ≈ 15 DU) were observed (Figure 3C). Binding of MRSA to phage probes and PBP2a antibody conjugated latex beads was confirmed using scanning electron microscopy investigations (Figure 3D).
When the phage spheroid biosensor was exposed to MSSA suspensions, a substantial decrease in the frequency (Δf ≈ -200 Hz) and an increase in the dissipation (ΔD ≈ 55 DU) were observed (Figure 3E). Following phage spheroid-MSSA binding interactions, assayed sensors were challenged with PBP2a antibody conjugated latex beads suspensions. Initially, the MSSA assayed biosensor showed short transients of increase in frequency and decrease in dissipation. After a few minutes of MSSA and PBP2a antibody conjugated latex beads interactions, frequency returned to post PBP2a antibody introduction levels, but the dissipative energy increased (Figure 3E). Binding of MSSA to phage probes was confirmed with scanning electron microscopy, but no binding between MSSA and PBP2a antibody conjugated latex beads was observed (Figure 3F). Ellipsometric thickness profile, 3D thickness map of lytic phages and staphylococcus bacteria are shown in Supplementary Figures S1 and S2.
Following contact of sensors with LB immobilized phages or spheroids to the suspension of 109 cells/ml of MRSA or MSSA, the bacteria were observed to bind phages or spheroids at density (ρ) of 9.1 x 107 (MRSA/intact phages), 7.9 x 107 (MRSA/spheroids), and 7.2 x 107 (MSSA/spheroids) cells/cm2. In tests using 5 different lytic phages and 2 host bacteria 27, researchers subjected phages immobilized by covalent binding method to 109 cells/ml bacteria, and determined the phage capture efficiency in a range of (2.5-8.9) x 105 cells/cm2. The phage capture efficiency presented in this work is ~100 times higher.
| Host | Strain | Strain details | Methicillin sensitivity | Phage sensitivity |
| S. aureus | 12600 | ATCC | S | + |
| S. aureus | 27690 | ATCC | S | + |
| S. aureus | 10292 | IA | S | + |
| S.aureus | 10378 | IA | S | + |
| S. aureus | 10497 | IA | S | + |
| S. aureus | 10686 | IA | S | + |
| S. aureus | MRSA 1 | AU | R | + |
| S.aureus | MRSA 2 | AU | R | + |
| S. aureus | MRSA 5 | AU | R | + |
| S. aureus | MRSA 13 | AU | R | + |
| S.aureus | MRSA 26 | AU | R | + |
| S. aureus | MRSA 34 | AU | R | + |
| S. aureus | MRSA 45 | AU | R | + |
| B. anthracis | Sterne | AU | NA | – |
| Salmonella typhimurium | LT2 | AU | NA | – |
| Shigella flexneri | unknown | AU | NA | – |
| Yersinia enterocolitica | unknown | AU | NA | – |
| Proteus mirabilis | unknown | AU | NA | – |
| Klebsiella pneumoniae | 13882 | AU | NA | – |
| Bacillus subtilis | 6051 | ATCC | NA | – |
Table 1. Phage 12600 sensitivity of bacterial strains. IA – isolated from animals; AU – bacterial culture collection of Auburn University; NA – not applicable; a – lytic activity of phage was defined by plaques formation, +, sensitive, -, not sensitive. Overnight cultures of tested strains were plated onto the plates with NZY agar. After the surface dried, a sample (10 μl) of 1011 PFU phage suspension was spotted on the surface of the plate. Plates were incubated at 37 °C for 18-24 hr. Lytic activity of the phages was detected by formation of plaques.
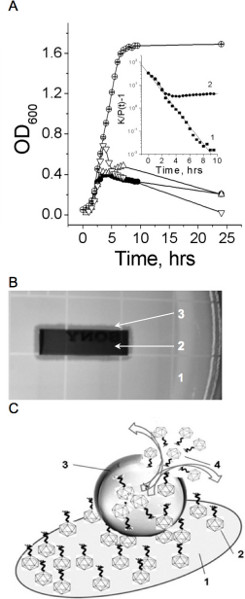
Figure 1. Infective properties of bound and free phage. A. Bacterial growth in the absence (empty circles) and presence (filled circles) of free phage, in the presence of phage bound to gold surface (top up triangles), and in the presence of phage bound to gold surface after 6 hours at 4 °C (top down triangles). Insert: The line (1) shows a linear fit of the no phage growth experimental data to the equation (4) (R=-0.99, p<0.0001). The line (2) shows a fit of the experimental data of bacterial growth at the free phage presence to the same equation. Bacterial growth constant (k) in the absence and presence of free phage, are equal 0.88 and 0.64, (-0.048, decline phase). Representative data of three independent experiments are shown in panel A. A mean relative error OD600 measurements did not exceed 5%. B. Phage immobilized to the gold surface is infective as indicated by the lysis zone around the gold piece. 1- the fragment the agar plate with bacteria; 2 – the gold piece with immobilized phage; 3 – the inhibition zone around the gold piece. C. Schematic representation of bacterium lysis by phage attached to the gold surface. 1 – gold-coated quartz piece, 2 – phage bound to the gold surface, 3 – bacterium anchored by bound phages, 4 – free phages have released by bursting infected bacterium. Used with permission from: Guntupalli, R., et al. 2012. Click here to view larger figure.
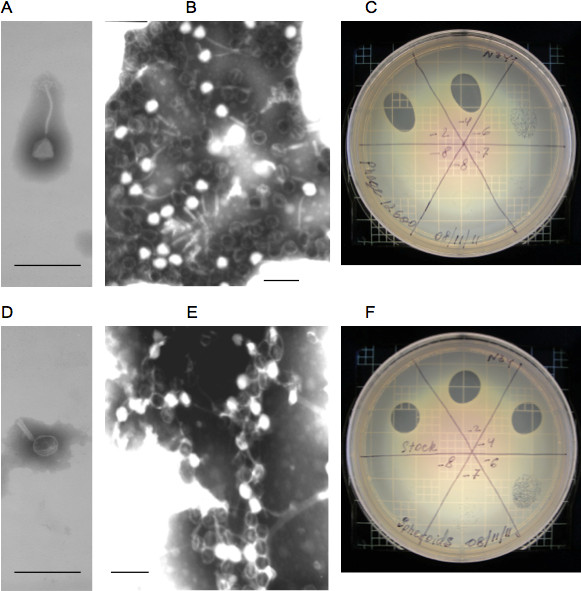
Figure 2. Properties of intact and modified phage. A and B – Transmission and scanning electron micrographs of intact phage, respectively; C – phage lytic activity on an agar plate with MRSA. D and E – Transmission and scanning electron micrographs of intact phage spheroids, respectively; F – phage spheroids lytic activity on an agar plate with MRSA. The mean activity of phage and spheroids are (7.4±1.5(SD)) x 1010 (N=4) and (7.5±1.0(SD)) x 1010 (N=7) PFU/ml. T-test: t = 0.15682, p = 0.87851. At the 0.05 level, the activities of phage and spheroids are NOT significantly different. Bars: A, B, D, and E: 200 nm. Used with permission from: Guntupalli, R., et al. 2012.
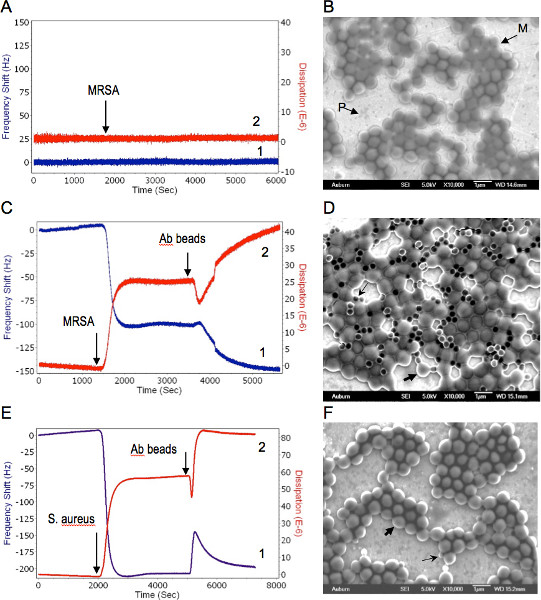
Figure 3. Combined QCM-D and EM analysis of phage-bacteria interactions. Bacteria were delivered to the sensor at concentration of 109 CFU/ml suspensions in water at a flow rate of 50 μl/min. 1, 2 represent changes in the resonance frequency and the energy dissipation, respectively. A. Phage coated QCM-D sensor response to MRSA. Arrow shows the MRSA delivery time to the sensor surface. B. Scanning electron micrograph of post assayed MRSA bound to lytic phage immobilized on the QCM sensor. M-MRSA, P-phages on the sensor surface. C. Successive responses of the phage spheroids coated QCM-D sensor to MRSA first and then to PBP antibody beads. Arrows indicate MRSA and PBP antibody delivery time to the sensor surface, respectively. D. Scanning electron micrograph of post assayed biosensor with phage spheroids, MRSA, and PBP antibody beads. Thick and thin arrows shows typical MRSA cell and antibody bead, respectively. E. Successive responses of the phage spheroids coated QCM-D sensor to S. aureus first and then to PBP antibody beads. Arrows indicate S. aureus and PBP antibody delivery time to the sensor surface, respectively. F. Scanning electron micrograph of post assayed biosensor with phage spheroids, S. aureus, and PBP antibody beads. Thick and thin arrows shows typical S. aureus cell and antibody bead, respectively. Used with permission from: Guntupalli, R., et al. 2012. Click here to view larger figure.
Supplementary Figures
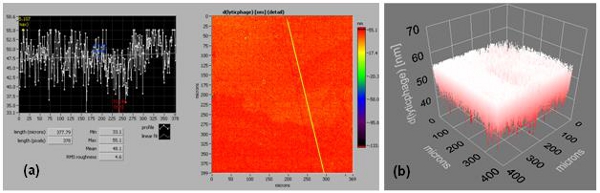
Figure S1. Supplementary Figure 1. (a) and (b) are an ellipsometric thickness profile and 3D thickness map of a lytic phage, respectively (effective refractive index = 1.05). Thickness profiles show mean, RMS roughness, minimum, and maximum thickness of the monolayer. A line across the thickness map was drawn to generate the thickness profile.
Figure S2. Supplementary Figure 2.
