For astrocyte specific expression of cyto-GCaMP3 in the striatum, we used adeno-associated virus (AAV) of the 5 serotype, and the GFAP GfaABC1D promoter (Figure 1A), which has been previously shown to drive robust GCaMP3 and reporter gene expression in hippocampal and cortical astrocytes8,14. Two weeks after virus microinjection into the mouse striatum, the mouse (~10 weeks old) was perfused and IHC was performed on thin brain sections to evaluate cyto-GCaMP3 expression in the striatum (Figure 1B). We detected cyto-GCaMP3 using GFP antibodies and also stained the slices with a known astrocyte marker, S100β. Cyto-GCaMP3 expression was only found in S100β positive cells. No expression was found in neurons visualized with NeuN (Figure 2A and 2B), suggesting astrocyte specific expression. Importantly, cyto-GCaMP3 expression was detected in ~60% of S100β positive cells (Figure 2C).
Astrocytes respond to brain insults such as injury, ischemia and infection by displaying changes that have become known as astrocyte reactivity, which represents a spectrum of potential changes from mild to severe15. We sought to evaluate if virus microinjection caused overt astrocyte reactivity. It was previously shown that reduced expression levels of Glutamine Synthetase (GS) were associated with the reactive astrocytes16,17. Therefore, GS expression in the striatum of WT and virus injected mice was compared. We found no significant changes in GS expression in striatal astrocytes following virus injection (Figure 3A – C), indicating no overt astrocyte reactivity using GS levels as a metric. This general approach could be expanded in future work to evaluate astrocyte reactivity using other markers of reactivity. However, note that GFAP levels may not be a suitable way to measure reactivity in striatal astrocytes, because GFAP is not expressed at detectable levels in most striatal astrocytes under basal conditions18. In summary, by using IHC we conclude cyto-GCaMP3 expression was robust and specific to astrocytes in the striatum, and that virus injection did not cause astrocyte reactivity as assessed by GS expression level changes.
We next prepared acute brain slices from virus injected mice to image [Ca2+]i in striatal astrocytes. Acute 300 μm thick sagittal or coronal slices were prepared, and after a period of recovery the slices were placed in a recording chamber on a confocal microscope for [Ca2+]i imaging using the 488 nm line of an Argon laser. Striatal astrocytes expressing cyto-GCaMP3 could easily be identified in most (usually all) of the striatal slices (~6 – 7 slices per mouse), as they showed visible green fluorescence in cells with clearly bushy morphology (Figure 4). A typical 5 min recording at 1 Hz from a representative field is shown in Figure 4A; at least 10 astrocytes displaying cyto-GCaMP3 could be imaged and are plotted in Figure 4A as Regions of Interest (ROI). Higher magnification imaging with an additional digital zoom of 2.0 – 3.0 was necessary to identify single astrocytes (Figure 4B). Under these circumstances, entire astrocyte territories were revealed by cyto-GCaMP3 expression, as shown in Figure 4B. Spontaneous Ca2+ signals were easily measured in the somata as well as in the branches and branchlets of astrocytes6.
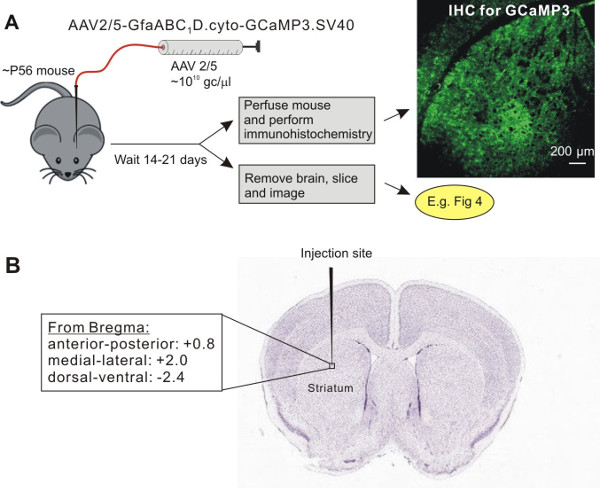
Figure 1: Viral delivery of GECIs (e.g. cyto-GCaMP3) by AAV2/5 into the adult mouse striatum. A. Schematic illustrates the protocol for AAV2/5 microinjections into the dorsolateral striatum. B. Shows the approximate position of the microinjection needle in relation to the striatum and the coordinates used for stereotaxic injections. The image of a Nissl-stained slice in panel B was downloaded from ALLEN BRAIN ATLAS.
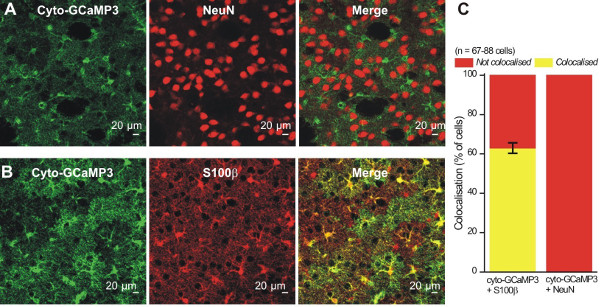
Figure 2: Cyto-GCaMP3 expression was specific to striatal astrocytes. A – B. Representative images showing that cyto-GCaMP3 expression colocalizes with a marker for astrocytes (S100β) (B), but not with a marker for neurons (NeuN) (A). C. Summary bar graph for colocalization experiments like those shown in A and B.
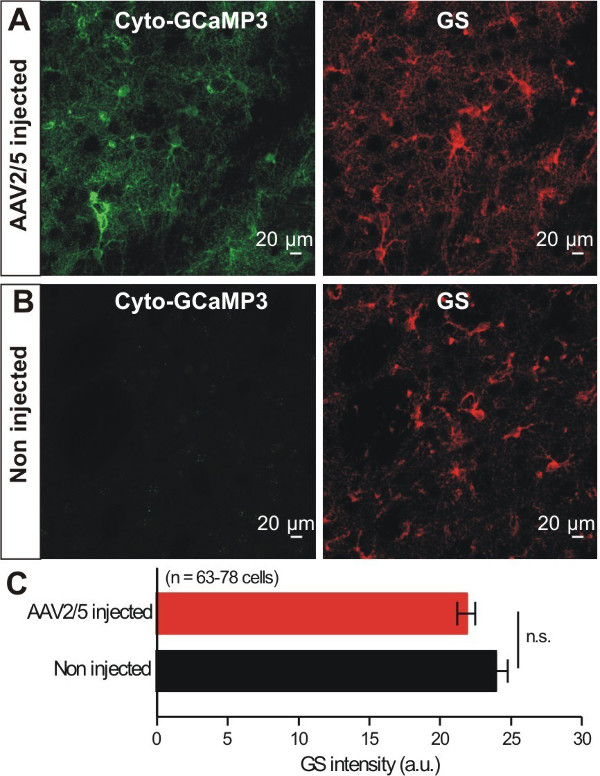
Figure 3: Cyto-GCaMP3 expression within striatal astrocytes did not cause changes in the expression of GS. A – B. Representative images of GCaMP3 and GS IHC from mice that had received AAV2/5 for GCaMP3. Images for control non injected mice are also shown. C. Bar graph summarizes results from experiments such as those in A and B (n.s. indicates not significant using an unpaired Student’s t test, p <0.05).
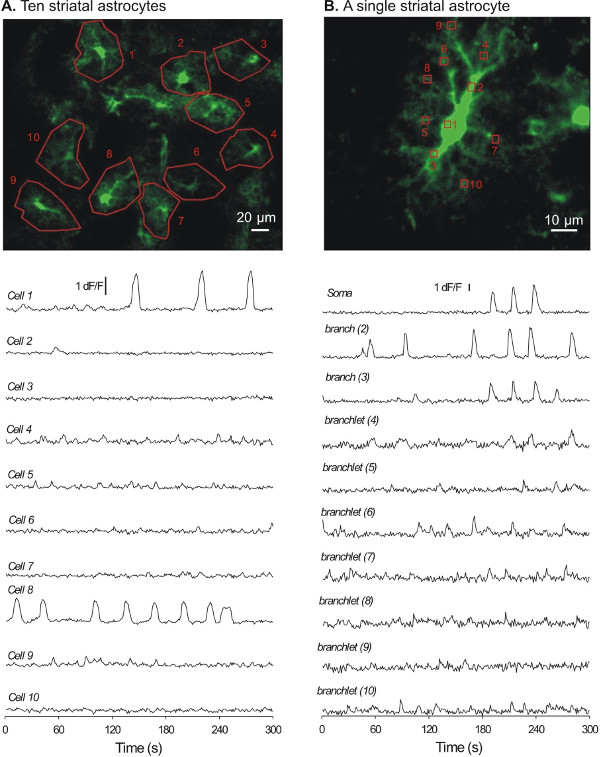
Figure 4: Representative examples of [Ca2+]i signals recorded from striatal astrocytes with cyto-GCaMP3. A. A flattened z-stack image showing 10 astrocytes (numbered 1 – 10).The traces below show [Ca2+]i signals recorded over 5 min from the 10 cells shown in A. B. A flattened and zoomed-in z-stack image for a single astrocyte.The traces below show [Ca2+]i signals recorded over 5 min for the single cell shown in B.
