In this report, we have demonstrated that conditions of varying iron availability have the capacity to modulate H. pylori Cag-T4SS pilus biogenesis at the host pathogen interface. When cultured in medium alone, H. pylori forms an average of 3 pili/cell. When H. pylori is grown in iron deplete conditions (using the synthetic chelator dipyridyl) that are sub-inhibitory to bacterial growth (Figure 1), the bacteria produce numerous Cag-T4SS pili (~7 pili/cell) when co-cultured with human gastric cells (Figure 2). Conversely, when an exogenous source of nutrient iron is present, the formation of Cag-T4SS pili is repressed (less than 1 pilus/cell in FE sample, less than 1 pilus/cell in FE+DIP sample) (Figure 2). Quantification of the pili at the host-pathogen interface reveals that in conditions of iron restriction, H. pylori exhibits an 2-fold increase in Cag-T4SS pili (p<0.0001) and the percent piliated cells increases by 11% (p<0.05) compared to cells grown in medium alone (Figure 3). Interestingly, the activity of the Cag-T4SS, as measured by host IL-8 secretion, is enhanced 107% under conditions of iron restriction and repressed 49% under conditions of excess iron availability compared to medium alone (p=0.03, and p<0.001, respectively) a result that supports the SEM data. However, pili dimensions remain consistent among all conditions of iron availability (Figure 4). Furthermore, iron availability does not change the biogenesis of pili on the surface of a cagE mutant, nor does it change this isogenic derivative’s ability to induce an IL-8 response from host cells, supporting the hypothesis that the structures demonstrated are, in fact, Cag-T4SS and not an additional unrelated surface feature.
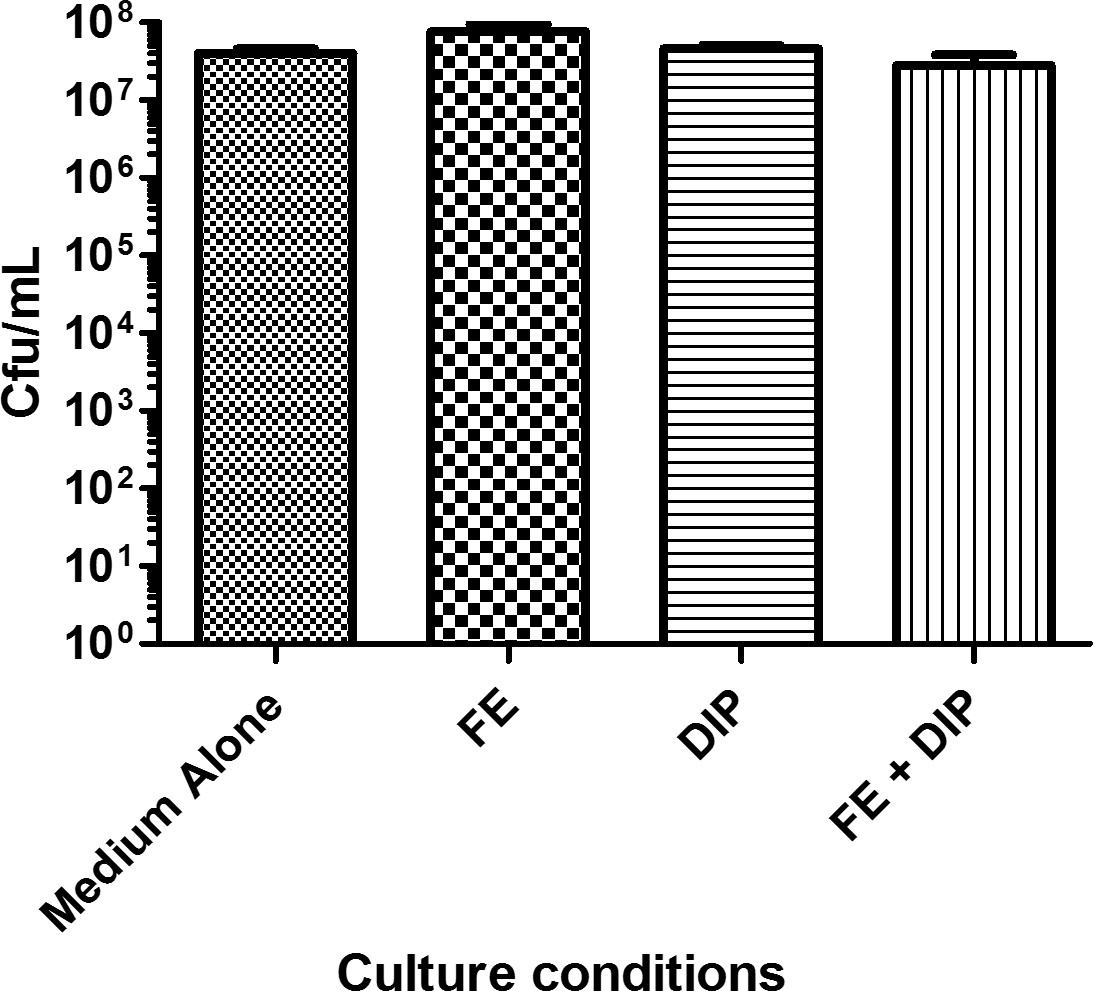
Figure 1: Bacterial viability determined in various conditions of iron availability. H. pylori was cultured in modified brucella broth alone (Medium Alone), or supplemented with 100 µM FeCl3 (FE), 200 µM dipyridyl (DIP), or 200 µM dipyridyl plus 250 µM FeCl3 (FE+DIP). Bacteria were diluted serially and plated onto bacteriological medium prior to incubation for 2 days at 37 °C in the presence of 5% CO2. Colonies were counted and colony forming units/mL (Cfu/ml) were calculated. Bars indicate mean of three independent experiments +/- SEM. Treatment with DIP, FE, or FE+DIP does not significantly alter bacterial viability.
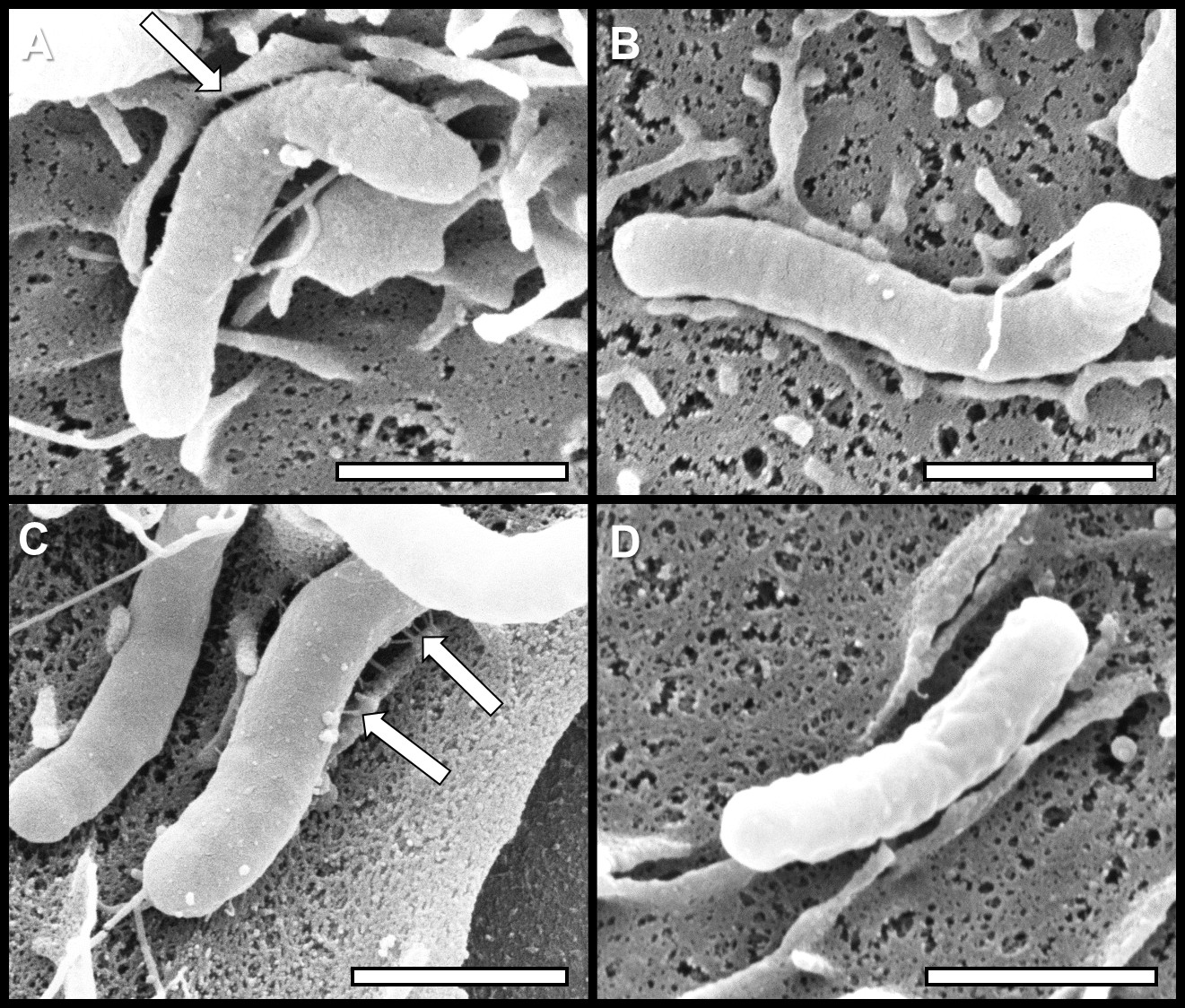
Figure 2: High resolution scanning electron microscopy analyses of H. pylori Cag-T4SS pili. Bacteria were grown in A) medium alone, B) medium supplemented with 100 µM FeCl3, C) medium supplemented with 200 µM dipyridyl, or D) medium supplemented with 200 µM dipyridyl plus 250 µM FeCl3 prior to co-culture with AGS human gastric cells. Arrows indicate Cag-T4SS pili formed at the host-pathogen interface. Samples were analyzed by high-resolution scanning electron microscopy to evaluate Cag-T4SS biogenesis. Conditions of low iron availability increase the prevalence of Cag-T4SS pilus formation, while conditions of excess available nutrient iron repress Cag-T4SS pilus formation.
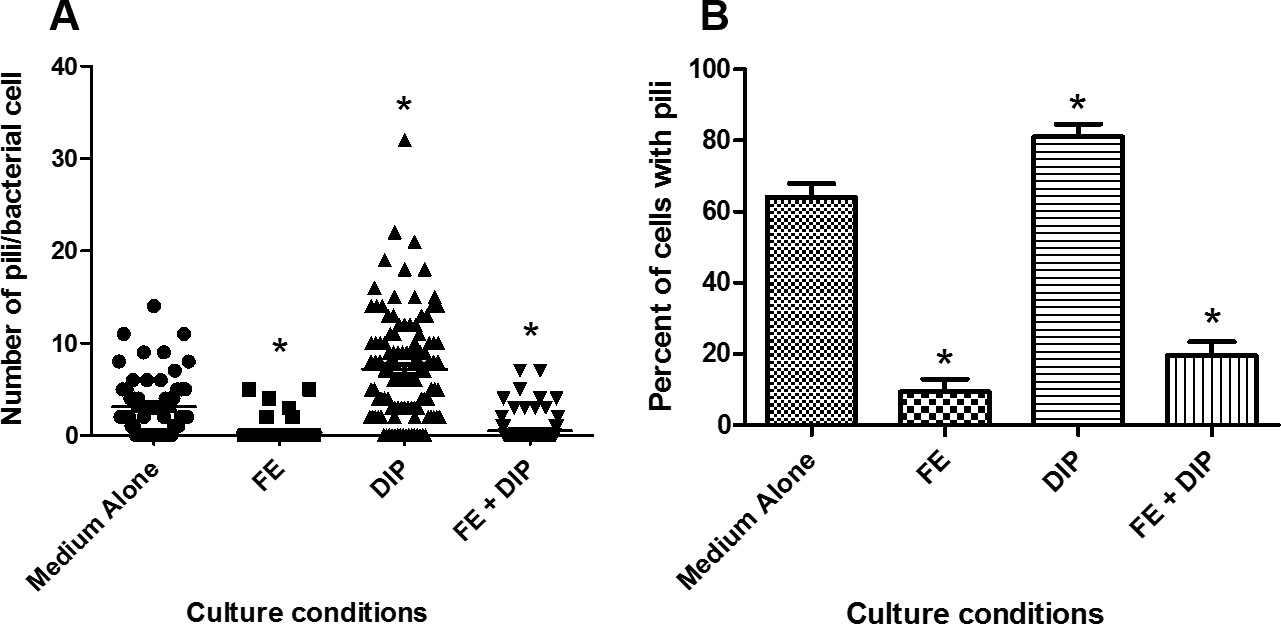
Figure 3: Quantification of pili/cell and percent piliated cells cultured in various conditions of iron availability. Bacteria were grown in modified brucella broth alone (Medium Alone), or medium supplemented with 100 µM FeCl3 (FE), 200 µM dipyridyl (DIP), or 200 µM dipyridyl plus 250 µM FeCl3 (FE+DIP) prior to co-culture with human gastric epithelial cells. Samples were analyzed by high-resolution scanning electron microscopy and pili were enumerated using ImageJ software. A) Scatterplot of pili per cell (*p<0.0001 compared to medium alone) and B) Bar graph depicting percent of piliated cells (*p<0.05 compared to medium alone) derived from 60-112 cells per culture condition derived from 3 separate biological experiments.
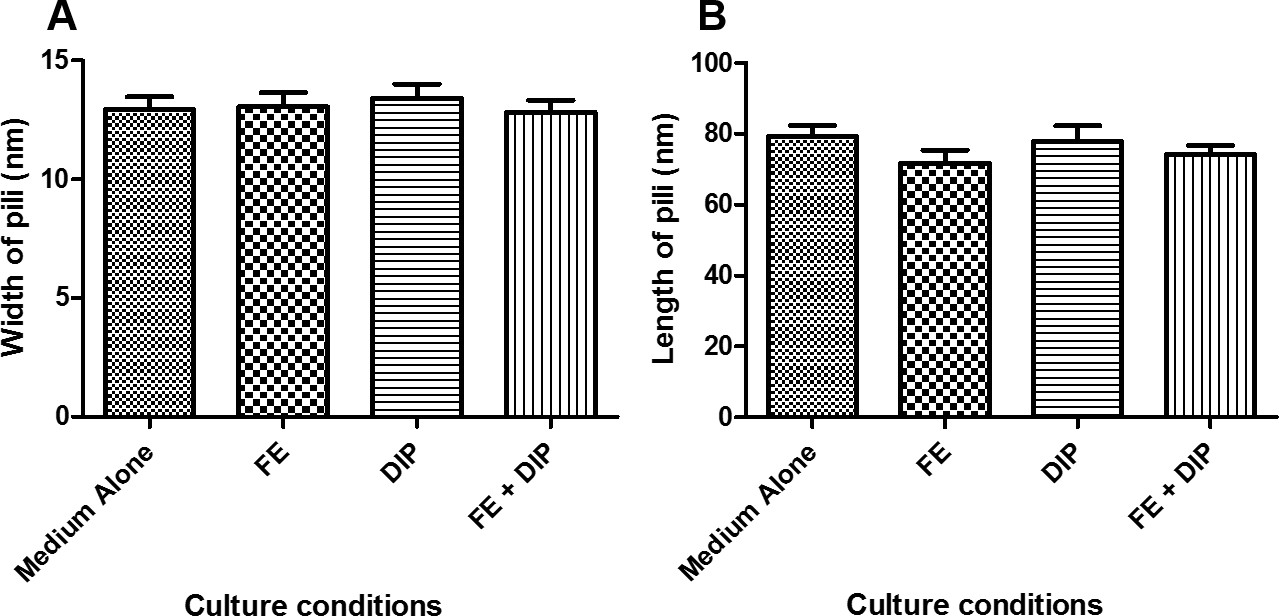
Figure 4: Pilus dimensions measured from cells cultured in various conditions of iron availability. Bacteria were grown in modified brucella broth alone (Medium Alone), or medium supplemented with 100 µM FeCl3 (FE), 200 µM dipyridyl (DIP), or 200 µM dipyridyl plus 250 µM FeCl3 (FE+DIP) prior to co-culture with human gastric epithelial cells. Samples were analyzed by high-resolution scanning electron microscopy and pili dimensions were measured using ImageJ software. A) Pilus width is an average of 13.1+/- 2.4 nm among all conditions. B) Pilus length is an average of 75.8 +/- 16.0 nm among all conditions. No significant difference in pilus dimensions are seen in the various conditions of iron availability.
Supplemental Figure 1: High resolution scanning electron microscopy analyses of H. pylori cagE mutant in various conditions of iron availability. Bacteria were grown in A) medium supplemented with 100 µM FeCl3, B) medium supplemented with 200 µM dipyridyl (DIP) or C) medium supplemented with 200 µM dipyridyl (DIP) plus 250 µM FeCl3 prior to co-culture with AGS human gastric cells. Samples were analyzed by high-resolution scanning electron microscopy to evaluate Cag-T4SS biogenesis. Conditions of low or high iron do not modulate any Cag-independent pilus biogenesis, supporting the hypothesis that the structures visualized in Figure 2 are Cag-T4SS pili.
Supplemental Figure 2: ELISA analysis of host IL-8 secretion in response to bacteria grown in varying conditions of iron availability. Wild-type (white bars) or cagE mutant (grey bars) bacteria were grown in modified brucella broth alone (Medium Alone), or medium supplemented with 100 µM FeCl3 (FE), 200 µM dipyridyl (DIP), or 200 µM dipyridyl plus 250 uM FeCl3 (FE+DIP) prior to co-culture with human gastric epithelial cells. Iron restriction results in increased IL-8, a host response that is dependent upon Cag-T4SS activity. Bars indicate mean +/- SEM, derived from 3 separate biological experiments. (*p<0.05 compared to PMSS1 grown in medium alone).
