A representative result of sex determination from 345 DNA lysates screening by PCR is shown in Figure 7 and summarized in Table 1.
As can be seen in Figure 7, the primers annealing temperature at 65 °C is the optimal condition in this PCR protocol generating similar intensity and predicated amplicon sizes (Figure 5) among different samples.
Two amplified products of SRY (400 bp) and ZFX (506 bp) are shown in the gel picture (Figure 7). Male embryos showed two DNA fragments with the correct size of the upper (400 bp) and lower (506 bp) bands. The intensity of these two bands was shown to be equal in most male individuals. All the female embryos only showed a single DNA fragment corresponding to the ZFX gene located in the X-chromosome.
DNA lysates obtained from the modified 50 mM NaOH can be used directly for PCR with a high success rate and accuracy. DNA lysates did not show any PCR reaction inhibition after screening 345 embryos. Only three individuals were not sexed and the percentage of female embryos was slightly higher than males (Table 1).
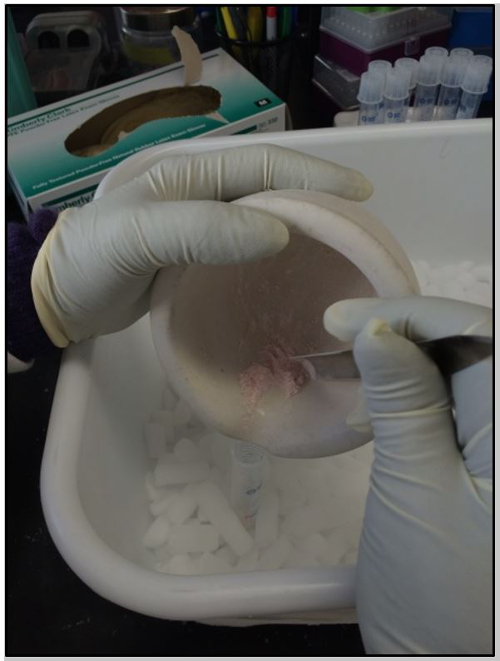
Figure 1: Embryo Powder Transfer. Transferring embryo powder to a 15 ml tube.
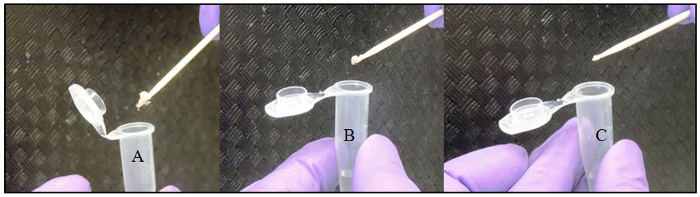
Figure 2: Collecting and Transferring Embryo Powder with a Toothpick. (A) An excessive amount of embryo powder adhering to the toothpick. (B) Correct amount of embryo powder adhering to the toothpick to be transferred to the alkaline lysis reagent solution. (C) An insufficient amount of embryo powder adhering to the toothpick.
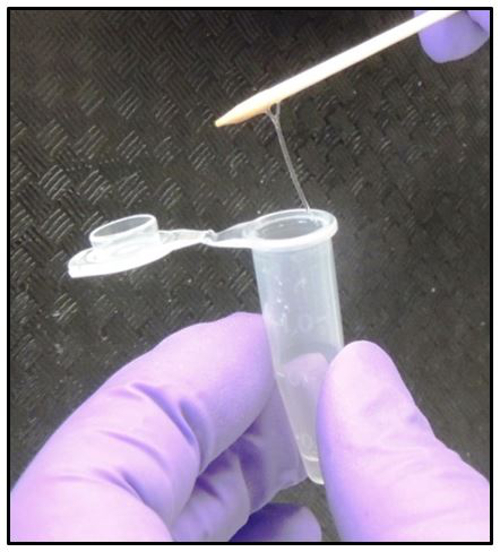
Figure 3: DNA Visualization Technique. Slowly pull up the toothpick, after adding the embryo powder, to determine if the embryo DNA has dissolved as a sticky gooey white transparent-like substance.
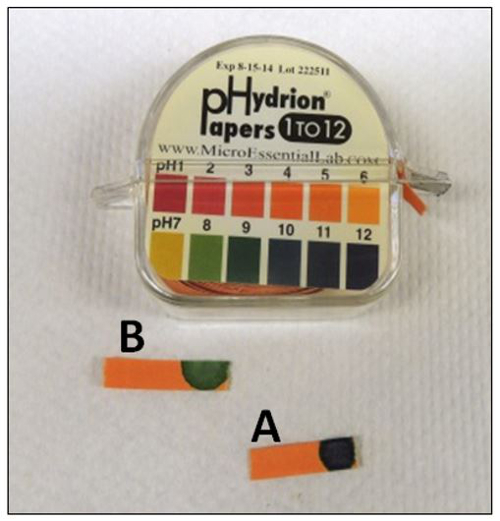
Figure 4: pH Confirmation. (A) pH of the sodium hydroxide (alkaline lysis reagent) solution is 12.0. (B) After addition neutralization buffer to the lysis buffer, color indication of pH paper is green and the pH should be around 8.0.
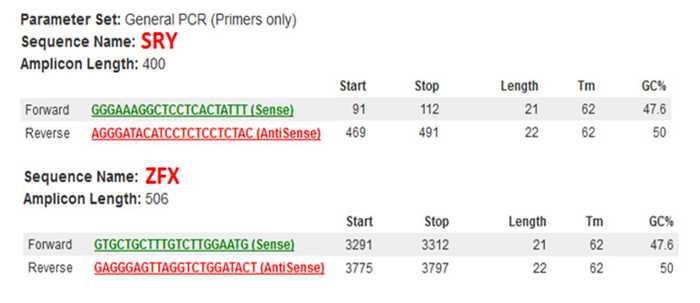
Figure 5: Sex-specific Primer Information. Sequence names, Sequences and the amplicon length obtained from the primer design tool. Please click here to view a larger version of this figure.

Figure 6: Mapping of Porcine Sex-specific Primers on Chromosome X and Y. (A) Location of the forward and reversed primers specified on the ZFX gene. (B) Location of the forward and reversed primers specified on the SRY gene. Please click here to view a larger version of this figure.
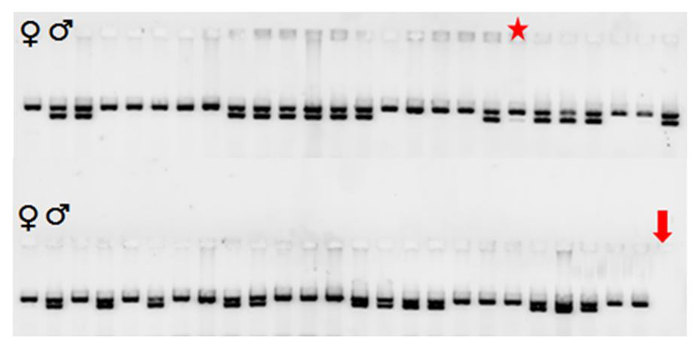
Figure 7: PCR Amplification of Porcine Sex-specific Genes from Day 30 Embryos. 2% TBE agarose gel stained with SYBR DNA gel stain with the known positive controls are indicated with male and female symbols. Two bands indicated as males and a single band as females. Red star indicated the possible sample contamination in the well with the appearance of a very faint lower band (400 bp) compared to upper band (506 bp) PCR product caused by SRY Y-specific primers. Red arrow indicates the no template PCR negative control. Please click here to view a larger version of this figure.
| Sexing PCR | Embryo Count | % |
| ♀ | 185 | 53.60% |
| ♂ | 157 | 45.50% |
| Unknown | 3 | 0.90% |
| Total | 345 | 100% |
Table 1: Percentage of Female, Male and Unknown after PCR Sexing among 345 Day 30 Embryos.
