Printing, Scanning & Data analyses
To ensure proper printing, it is vital to have the correct alignment of the spotted grid within the teflon mask, which delineates each array on the slide. During printing, due to the nature of the teflon coating, spots cannot be seen by the naked eye on the MPX slides. Attention is paid then to the appearance of the pre-spots on the poly-L-Lysine coated slides. Directly after printing, each slide should be checked by eye for the presence of each spotted compound that is visible due to the buffer's salts dried in the spot. Arrays are inspected for correct alignment of spots within the Teflon borders and correct number of spotted features.
Slides are scanned in an iterative process of lower to higher scan power to avoid photo bleaching and allowing the comparison of higher avidity to lower avidity binding proteins, which have higher and lower output signals, respectively. Following scanning, the fluorescent intensity of each spot in the output images are measured using an imaging program. Each array is overlayed with a grid file that matches the number of features printed on the array surface (Figure 1D–G). Using printed fluorescent dyes, the borders of the printed glycans are defined and a mask that is integrated with the identity of the printed sample lassoes each spot. The imaging mask lasso will record all aspects of the imaged spot (size, signal intensity, coordinates in the grid, etc.) and output this information to a tab-delimited text file. Using tabulation software (spreadsheet), the samples are sorted by identity and the mean signal intensity minus background is averaged over 4 of the 6 spots for each sample, leaving out the highest and lowest signal as outliers. A line graph of average mean signal minus background is plotted against the eight protein concentrations applied to the array and contains one line for each sample. The final output graph is smoothed using a non-linear regression calculation (Figure 1I).
Glycan binding proteins
The PAA-Array is used to assess receptor-binding specificity of influenza A virus hemagglutinins. An additional feature of our array, not present in the analogous plate assay, is the inclusion of non-sialylated controls in the same micro-well. To assess that printed compounds are present after printing, commonly used plant lectins with known specificity were used. ECA binds to terminal galactose linked β1-4 to N-acetyl-glucosamine (Galβ1-4GlcNAc or LacNAc) and will not bind if the terminal galactose is capped with sialic acid. ECA only detects the non-sialylated glycans on our miniaturized glycan array (Figure 2A). The lack of binding to any of the sialylated glycan samples is also indicative that these are fully capped with terminal sialic acid. For α2-6 linked sialic acid, SNA was used as a lectin control (Figure 2B). As expected, SNA only binds to α2-6 linked sialic acids, but with notable differences in affinity for PAA-linked non-PAA-linked di-LacNAc repeats. This difference reflects the sensitivity of PAA-linked glycans and allows the discrimination of proteins that may bind with low avidity. For α2-3 linked sialic acid, an H5 hemagglutinin derived from A/Vietnam/1203/04 virus that is known to recognize α2-3 linked sialic acids only was used 6. The binding profile of the H5 HA indeed shows specificity for α2-3 linked sialic acids, with a higher avidity for the PAA-linked structures (Figure 2C). Finally, the H1 hemagglutinin from a human seasonal H1N1 strain was used that, as expected, only bound to α2-6 linked sialic acid containing structures (Figure 2D).
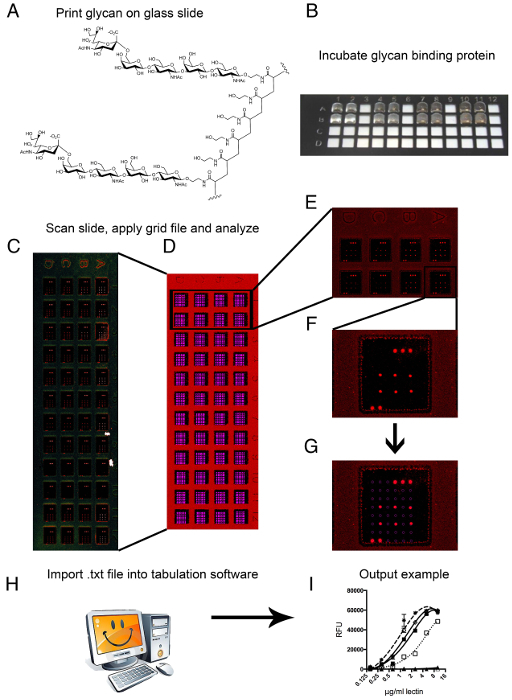
Figure 1: Printing, scanning and image analyzes. (A) PAA-conjugated 6SLNLN is shown as a representative glycan structure that is printed on the glass slides. (B) Incubating a glycan binding protein on the multi-well glycan array is shown with an 8 µl volume that creates a droplet on the array surface. (C) Following incubation of the glycan binding proteins on the array and scanning in a confocal fluorescent slide scanner, a representative image is obtained. (D) The image is overlayed with a grid and, using grid markers for proper alignment, the single spots can be analyzed. (E) A close up of a complete set of 8 arrays, in which a single glycan binding protein was analyzed using eight 1:1 dilutions. (F) A single array, in a single well is represented; the array is demarked by the grid markers in the top right (3 spots) and bottom left (2 spots), this glycan binding protein binds a single compound on the array as a replicate of six is clearly visible. (G) The grid is shown with lassos encircling specific individual spots. (H) Imaging software calculates signal values from the image file and outputs a tab-delimited data file. (I) The data can be tabulated in spreadsheet or stastical software to create a representative output graph. Please click here to view a larger version of this figure.
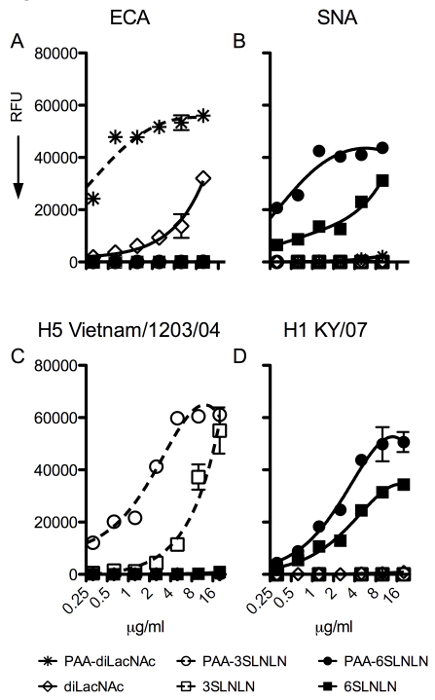
Figure 2: Output of plant lectins (ECA, SNA) and IAV hemagglutinins with different specificities (H5 from A/Vietnam/1203/04, H1 from A/Kentucky/07). (A) ECA binds terminal LacNAc or Galb1-4GlcNAc specifically and detects the presence of the non-sialylated compounds, used as controls for IAV hemagglutinins. (B) SNA recognizes only glycans bearing a terminal α2-6 sialic acid. (C) The recombinant hemagglutinin of the H5N1 (A/Vietnam/1203/04) strain binds to α2-3 sialic acids and provides an avian type receptor binding profile. (D) The recombinant hemagglutinin of a human seasonal H1N1 (A/Kentucky/07) binds to human-type receptors only. Fluorescent signal intensity was measured for each spot, and mean intensity minus mean background was calculated using spreadsheet program. For each glycan, the mean signal intensity is calculated from 6 replicates spots. The highest and lowest signals of the 6 replicates are removed and the remaining 4 replicates are used to calculate the mean signal, standard deviation (SD), and standard error measurement (SEM). The graphs represent the averaged mean signal minus background for each sample and error bars are the SEM value. Please click here to view a larger version of this figure.
